

El circovirus porcino tipo 2 (PCV2) es uno de los virus más extendidos que causa infecciones económicamente relevantes en los cerdos.
Tras su primera identificación a finales de los años 90, el PCV2 se ha vinculado a diversas manifestaciones de la enfermedad (Allan 1998; Rosell et al. 2000; Kim, Chung, and Chae 2003; Madson and Opriessnig 2011).
Síndrome Multisistémico de Desmedro Posdestete (PMWS)
Síndrome de dermatitis y nefropatía porcina (PDNS)
Complejo respiratorio porcino (PRDC)
Insuficiencia reproductiva

Todas estas enfermedades se denominan colectivamente enfermedades asociadas al circovirus porcino (PCVAD) (Opriessnig, Meng, and Halbur 2007).


Esto demostró que el PCV2 circulaba años antes de su identificación.
El PCV2 pertenece al género Circovirus (familia Circoviridae) y se trata de un virus de ADN, icosaédrico y sin envoltura, con un diámetro de 17 nm.
![]()
![]()
ORF1 Codifica dos proteínas asociadas a la replicación (rep y rep0) que son esenciales para la multiplicación del genoma del PCV2 pero que están ausentes en el virión ensamblado.
ORF2 Codifica una proteína de cápside viral (cap), la única proteína estructural.

La proteína de la cápside (cap) determina la antigenicidad y virulencia del PCV2 (Hamel, Lin, and Nayar 1998; Nawagitgul et al. 2000; Lefebvre et al. 2008; Saha et al. 2012b)., y la mutación de uno o dos de sus aminoácidos puede afectar a esta virulencia y patogenicidad (Huang et al. 2011; Saha et al. 2012a).
Por otra parte, el ORF2 por sí solo puede servir como marcador filogenético y epidemiológico para el análisis comparativo del PCV2 (Olvera, Cortey, and Segalés 2007).
![]()

PCV2a & PCV2b
PCV2a y PCV2b se han documentado en todo el mundo (Segales et al. 2008). PCV2a fue el genotipo más prevalente hasta aproximadamente 2003, mientras que PCV2b se convirtió en el predominante después de 2004 (Segales, Kekarainen y Cortey 2013).
Esto indica un evidente cambio de genotipo de PCV2a a PCV2b que se describió en varios países (Carman et al. 2006; Cheung et al. 2007; Dupont et al. 2008; Timmusk et al. 2008; Wiederkehr et al. 2009; Cortey et al. 2011; Segales, Kekarainen, y Cortey 2013).
![]()

PCV2c
El PCV2c se detectó inicialmente en muestras archivadas de suero de cerdo en Dinamarca, y se ha encontrado recientemente en un cerdo salvaje en Brasil (Dupont et al. 2008; Franzo et al. 2015). Sin embargo, es probable que este genotipo sea de menor importancia.
PCV2d
El origen de las cepas PCV2d se remonta a 1998 en Suiza y actualmente está muy difundido en muchos países, lo que sugiere un cambio de genotipo de PCV2b a PCV2d (Wei et al. 2013; Xiao, Halbur y Opriessnig 2015; Kwon et al. 2017; Ramos et al. 2017; Thangthamniyom et al. 2017).
PCV2d se divide en dos subclases PCV2d-1 y PCV2d-2 (Xiao, Halbur y Opriessnig 2015).
![]()



El quinto genotipo, PCV2e, fue descrito por primera vez en cerdos mexicanos y luego en los EE.UU. en un estudio retrospectivo, y la secuencia más temprana se remonta a 2006 (Harmon et al. 2015; Davies et al. 2016).
PCV2f
El último genotipo, PCV2f, fue identificado por primera vez en China y también está presente en Croacia, India e Indonesia (Bao et al., 2018)


En Bélgica, la presencia de PCV2 se demostró en 1969 en base a un análisis serológico retrospectivo para la detección de anticuerpos de PCV2 en poblaciones de cerdos domésticos y silvestres (Sánchez, Nauwynck y Pensaert 2001a).
Nunca se han descrito grandes brotes de PMWS en Bélgica (Labarque et al. 2000; Meerts et al. 2004).
Se cree que el uso de los verracos Pietrain para la obtención de cerdos de engorde híbridos constituye la base de esta baja susceptibilidad.
Trabajos recientes han demostrado una asimilación y desintegración más eficiente del PCV2 por parte de los monocitos de la raza pura Pietrain con respecto a las razas puras Landrace y Large White, lo que podría explicar, en parte, esta mejor resistencia (Wei et al. 2018).
Hasta ahora, solo se han aislado cuatro cepas belgas a partir de lechones afectados por el PMWS:
Las cepas 1206 y VC2002-K39 fueron identificadas como PCV2b
La cepa VC2002-K2 como PCV2d-1
La cepa 4D4 como una cepa recombinante de VC2002-K2 y VC2002-K39
(Meerts et al. 2004, 2005; Lefebvre et al. 2008, 2009; Saha et al. 2014; Xiao, Halbur y Opriessnig 2015).

A pesar de los continuos informes sobre la aparición de nuevas cepas y los cambios del genotipo a nivel mundial, se disponía de poca información sobre la epidemiología molecular y la evolución del PCV2 en Bélgica.


Examinar la prevalencia y la evolución genética del PCV2 en Bélgica desde 2009 hasta 2018.
Encontrar la tendencia evolutiva del PCV2 mediante el análisis de las diferencias de los aminoácidos.
MATERIAL & MÉTODOS
Entre 2009 y 2018 se tomaron 319 muestras (pulmones, riñones, bazo, ganglios linfáticos, suero o una mezcla de órganos) de cerdos con signos clínicos de PCVAD en diferentes regiones de Bélgica.
El análisis de las muestras consistió en:

La titulación de PCV2 se realizó en las células PK-15 (Sánchez y otros, 2001b), expresándose como log10TCID50/g de tejido, clasificándose las muestras en tres grupos:
Títulos altos: 4,5 log10TCID50/g
Títulos medios: 3-4,5 log10TCID50/g
Títulos bajos: 3 log10TCID50/g

Se extrajo ADN a partir de las muestras de tejido homogeneizados o suero, examinando los productos de la amplificación en gel de agarosa. Los amplicones fueron secuenciados y analizados para su comparación.
En el análisis filogenético se incluyeron 43 secuencias de nucleótidos ORF2 obtenidas en este estudio y 29 secuencias ORF2 de PCV2 publicadas y disponibles en el GenBank.
Se alinearon todas las secuencias y se realizó un análisis de similitud de las secuencias de nucleótidos y de aminoácidos para crear un árbol filogenético.
MAPEO TRIDIMENSIONAL (3D) DE LOS AMINOÁCIDOS EN LA PROTEÍNA CAP 
RESULTADOS
PORCENTAJE DE CASOS POSITIVOS DE PCV2 EN BÉLGICA (2009-2018) 


El 8% de las muestras contenía grandes cantidades de PCV2 infectante (4,5 log10TCID50/g)
El 1% de las muestras contenía cantidades moderadas de PCV2 infectante (3-4,5 log10TCID50/g)
El 7% de las muestras contenía cantidades bajas de PCV2 infectante (3 log10TCID50/g)
El 84% de las muestras dieron negativo para PCV2
SECUENCIACIÓN Y ANÁLISIS FILOGENÉTICO DE LAS SECUENCIAS ORF2 DE PCV2 
Un total de 43 muestras fueron sometidas a la extracción de ADN y a la amplificación por PCR del gen ORF2. El árbol filogenético mostró que las 43 secuencias de PCV2 obtenidas podían clasificarse en tres genotipos distintos (Figura 1):
3 de los 43 (7%) aislados de PCV2 pertenecían al genotipo 2a
31 de los 43 (72%) aislados de PCV2 pertenecían al genotipo 2b
9 de los 43 (21%) aislados de PCV2 pertenecían al genotipo 2d
Los genotipos PCV2c, PCV2e y PCV2f no fueron detectados en el conjunto de muestras
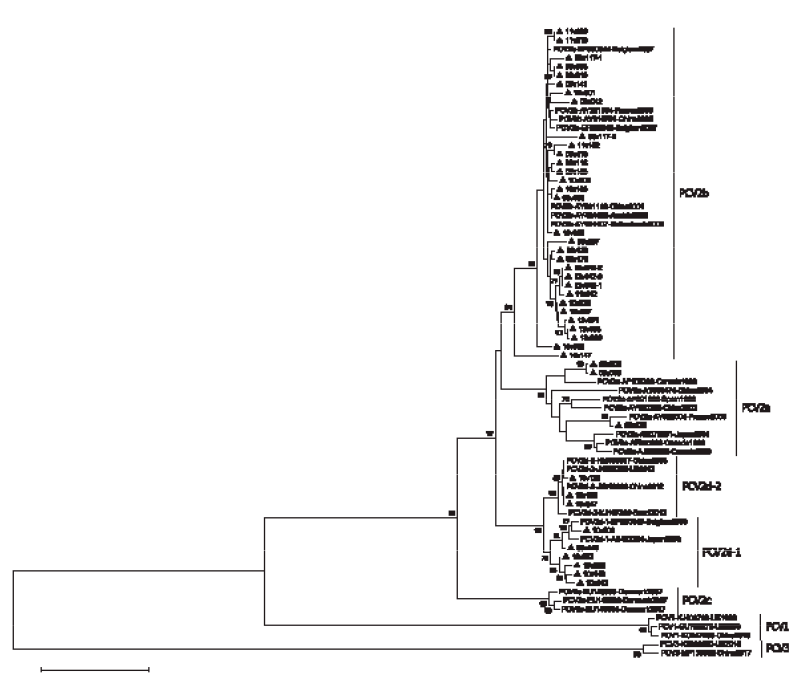


Las tres muestras del genotipo PCV2a fueron recogidas en 2009. El PCV2b fue el genotipo más común encontrado entre 2009 y 2013, con una diversidad genética relativamente limitada. El PCV2d formó un grupo independiente que se subdividió en dos subclases separadas: PCV2d-1 y PCV2d-2.
Curiosamente, los aislados de PCV2d-1 se encontraron en 2009 y en 2010, mientras que en 2018 todas las secuencias de PCV2d se agruparon en el subtipo PCV2d-2.
La longitud del ORF2 de PCV2a y PCV2b era de 702 pb, mientras que la de PCV2d era de 705 pb, lo que resultaba en un aminoácido adicional, la lisina, en la posición 30 de la proteína de la cápside.



El gen ORF2 de 6 aislados de PCV2d-1 demostró tener una similitud de secuencia de nucleótidos del 97,9-99,4% y una similitud de secuencia de aminoácidos del 97,0-99,1%, mientras que 3 secuencias de PCV2d-2 tuvieron unas similitudes de 97,7-100% y 97,0-100%.

Se estimó la presión selectiva sobre la proteína de la cápside en los aislados de PCV2 belgas. Al estimar la diferencia entre las tasas de sustitución no sinónimas y sinónimas (dN-dS), el gen de la cápside resultó negativo (1,575), lo que indica que la cápside de la PCV2 belga había evolucionado bajo una selección purificadora.
Sin embargo, se identificaron tres codones (posiciones 59, 131 y 191) bajo selección positiva:
Posición 59:Aminoácido crítico que forma parte del epítopo de neutralización (Huang et al. 2011).
Posición 131: Importante para determinar la actividad de neutralización (Saha et al. 2012a).
Posición 191:La mutación en esta posición puede afectar a la capacidad de crecimiento de PCV2 tanto in vitro como in vivo (Fenaux et al. 2004a).
EL ANÁLISIS DE AMINOÁCIDOS DE LAS SECUENCIAS DE CAP DE PCV2
La comparación de los aminoácidos de los 43 aislados de PCV2 y las 21 cepas de referencia revelaron una mayor diversidad en:
![]()
![]()
Además, ciertos aminoácidos solo aparecen en cepas de uno, pero no en los otros dos genotipos.
Por ejemplo, 10 residuos de aminoácidos (75N/K, 76L, 86T, 88K, 89I, 91I, 123I/V, 136Q, 190S/T, y 232N/K) solo se encontraron en grupos de aislados de PCV2a; 15 (11K/R, 29F/L, 82K/P, 85H/G, 89L/R, 112P/T, 141S/Y, 169A/S, 188H/Q, 190A, 206V, 210D/G/E, 215G/I/V, 217L/M, y 222T/R) en aislados de PCV2b, y 12 (10G/R, 53I, 59A/K/R, 68N, 130I/V, 134N, 169W/R/G, 188P/Q, 200P/T, 207H/Y, 215I, y 216A/T) en los grupos de PCV2d.
Otras mutaciones estaban presentes en todas o, al menos, en la mayoría de las cepas del genotipo, como la 57V/I en las cepas PCV2b, con V presente en dos de las 31 cepas PCV2b aisladas y una en las otras 29 cepas.


Los aminoácidos en estos importantes puntos de mutación fueron mapeados en la correspondiente estructura tridimensional de la proteína cap de PCV2a (cepa Fh17), PCV2b (cepa 1206) y PCV2d (cepa K2).
ESTRUCTURA INTERNA DE LA PROTEÍNA CAP
En la cara interna de la proteína de la cápside, donde interactúa con el genoma viral, se encontraron tres mutaciones en las posiciones 53 (F-F-I), 121 (SS-T) y 215 (V-V-I) de la lámina β en los tres genotipos (PCV2a-PCV2b-PCV2d).
Estas mutaciones son conservadas y es menos probable que induzcan algún cambio, ya que F, I y V pertenecen a aminoácidos hidrofóbicos, y S y T a aminoácidos polares.

ESTRUCTURA EXTERNA DE LA PROTEÍNA CAP
Curiosamente, la mayoría de las mutaciones en los aminoácidos se encontraron en el exterior de la proteína cap, donde interactúa con el entorno.
PARTE SUPERIOR DE LA PROTEÍNA CAP
En la parte superior de la proteína de la cápside se encontraron mutaciones en aminoácidos en las posiciones 59, 206 y 63 que estaban muy cerca entre sí y formaban una gran superficie (A/K/S para PCV2a, R/I/R para PCV2b y A/K/R para PCV2d).
Es evidente que esta área se hizo más básica con la evolución de PCV2a a PCV2b y PCV2d, debido a la formación de un grupo de aminoácidos polares básicos.
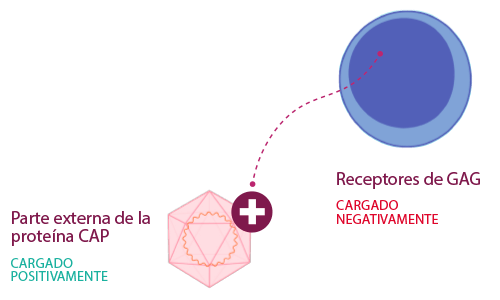


Otra gran superficie donde se encontraron mutaciones fue la formada por aminoácidos en las posiciones 57, 68 y 134, V/A/T para PCV2a, I/A/T para PCV2b, y V/N/N para PCV2d.
En comparación con PCV2a y PCV2b, esta zona de PCV2d aumentó su polaridad al añadir un segundo aminoácido polar (NN) en lugar de un aminoácido no polar (A, I o V), creando un mejor ambiente para la unión de los receptores a través de enlaces de hidrógeno.
Todo esto sugiere que, con las mutaciones de los aminoácidos en la parte superior de la proteína cap, el PCV2 ha evolucionado para lograr una unión más eficiente con los GAG.
PARTE INFERIOR DE LA PROTEÍNA CAP
En la parte inferior de la proteína cap había tres grandes áreas de superficie que estaban formadas por grupos de aminoácidos mutados en tres genotipos:
El primero (V/D/S para PCV2a, I/N/A para PCV2b, y L/N/T para PCV2d) estaba formado por aminoácidos en las posiciones 80, 77 y 190.
El segundo (V/S/I para PCV2a, L/S/V para PCV2b, y L/T/V para PCV2d) estaba formado por aminoácidos en las posiciones 80, 90 y 91.
El último (T/K/I para PCV2a, S/P/R para PCV2b, y S/P/L para PCV2d) estaba formado por aminoácidos en las posiciones 86, 88 y 89.
En general, las mutaciones en las dos primeras áreas estaban conservadas, ya que los aminoácidos mutados fueron reemplazados por otros con propiedades bioquímicas similares. Sin embargo, las mutaciones en la última área dieron lugar a una pérdida del aminoácido polar básico cargado positivamente.
Esto significa que hubo menos cambios en la parte inferior de la proteína cap.
Por otro lado, la superficie del aminoácido polar básico K en la posición 234 estaba muy cerca del aminoácido polar N en la posición 232 para PCV2d.
Se dedujo que el aminoácido polar N en la posición 232 podía extenderse más allá de la superficie de la cápside, proyectando así el aminoácido K con carga positiva vecino hacia el entorno exterior para buscar/ mejorar la interacción inicial inespecífica del virus con el heparán/condroitín sulfato con carga negativa de la membrana celular durante la infección por PCV2.


En conclusión, los patrones de aminoácidos eran multivariantes en las secuencias de la proteína cap en estos 43 aislados belgas de PCV2.
Desde el punto de vista evolutivo, la mayoría de los aminoácidos mutados se encontraron en la superficie exterior de la proteína cap. Las partes superior y posterior de la proteína cap contaban con grupos de aminoácidos más básicos y la parte inferior estaba más conservada, pero algo menos básica. Los aminoácidos de la parte interna se encontraban conservados para interactuar con el genoma viral y mantener la estabilidad del virión PCV2.
*Aminoacidos: N: Asparagina; K: Lisina; L: Leucina; T: Treonina; V: Valina; I: Isoleucina; Q: Glutamina; S: Serina; R: Arginina; F: Fenilalanina; P: Prolina; H: Histidina; G: Glicina; Y: Tirosina; A: Alanina; D: Ácido aspártico; M: Metionina; W: Triptófano; E: Ácido glutámico; C: Cisteína
CONCLUSIONES
Este es el primer estudio de caracterización molecular de las cepas belgas de PCV2.
Durante el período 2003-2004, se produjo un cambio global del genotipo 2a hacia el 2b. Esto concuerda con el predominio de PCV2b de 2009 a 2013 en Bélgica. En 2018, el PCV2d, especialmente el PCV2d-2, era el único subtipo que circulaba en Bélgica en nuestro estudio.
En varios estudios se ha sugerido la emergencia y propagación de PCV2d-2, que parece estar reemplazando a PCV2b (Xiao, Halbur y Opriessnig 2015; Franzo et al. 2016).
La aparición de PCV2d-1 coincidió con la propagación de PCV2b, lo que indica que PCV2b y PCV2d se bifurcaron pronto y evolucionaron de forma independiente en la población porcina (Xiao, Halbur y Opriessnig 2015).

La presión selectiva juega un papel crítico en la biología evolutiva al dar forma a la variación genética. La elevada tasa de mutación del PCV2, que se aproxima más a la de un virus de ARN de cadena simple, puede aumentar la frecuencia de la selección, así como la rápida aparición de nuevos genotipos del PCV2.
La vacunación amplia utilizada para controlar numerosas PCVAD, podría contribuir a la alteración de las propiedades antigénicas de PCV2. Investigar el papel de la vacunación en la evolución de la PCV2 es extremadamente difícil debido a la dificultad de reclutar un gran número de granjas donde no se haya realizado una vacunación a pesar de la circulación de la PCV2.
Sin embargo, es interesante observar que el primer cambio de genotipo de PCV2a a PCV2b durante el período 2003-2004 es anterior a la vacunación frente a PCV2 utilizando vacunas comerciales tipo PCV2a. Este cambio/evolución del genotipo probablemente estuvo impulsado por la inmunidad frente a la infección presente en todas las granjas.
Aunque el actual cambio de genotipo de PCV2b a PCV2d se produce en el contexto de una amplia vacunación con las vacunas tipo PCV2a, las infecciones de PCV2 siguen produciéndose e inducen una fuerte inmunidad de la población. La vacunación no ha reducido la variabilidad de las cepas de PCV2 en los rebaños vacunados (Reiner, Hofmeister y Willems 2015). Como resultado, es probable que la inmunidad de la población frente a los virus PCV2 de campo circulantes sea la principal fuerza impulsora de su evolución genotípica.
Dado que la proteína de la cápside de PCV2 es inmunogénica, las mutaciones en la proteína de la cápside pueden explicar la dirección de la evolución genotípica de PCV2.
En este estudio, los aminoácidos de la parte interna de la proteína cap, donde interactúa con el genoma viral, estaban conservados para mantener la estabilidad del virión, y la mayoría de las mutaciones de aminoácidos se encontraron en la superficie, donde interactúa con el entorno.
Es probable que la proteína de la cápside haya mutado para escapar de la inmunidad del huésped, evolucionando para unirse más fuertemente a sus receptores GAG.
Asimismo, PCV2d parece haber formado una parte extensible en la proteína cap que mejora la eficiencia de unión del virus a su célula diana.
La comparación de las secuencias ORF2 completas de los 43 aislados belgas de PCV2 reveló una homología de los nucleótidos del 89,7-100% y una similitud de las secuencias de aminoácidos del 88,5-100%.
Se encontró una mayor diversidad en las regiones 53-91, 121-136 y 169-217 y en las posiciones 151, 232 y 234 y estas regiones contienen epítopos inmunorrelevantes (Mahe et al. 2000). Así, mutaciones frecuentes pueden cambiar las características antigénicas del PCV2 y ayudar al virus a evadir la respuesta inmunitaria, promoviendo así la propagación del virus en condiciones de campo:
La posición 59 es un aminoácido crítico que forma parte del epítopo de neutralización (Huang et al. 2011).
Las posiciones 131 y 190 son importantes para determinar la actividad neutralizante (Saha et al. 2012a).
La región 163-180 se ha sugerido como epítopo señuelo que da lugar a la producción de anticuerpos no neutralizantes frente a PCV2 (Trible et al. 2011, 2012).
La posición 233es crítica para mantener la unión efectiva entre el epítopo y el anticuerpo, que se conserva entre todos los genotipos del estudio (Shang et al. 2009).



BIBLIOGRAFÍA
Allan, G. M. (1998) ‘Isolation of Porcine Circovirus-Like Viruses from Pigs with a Wasting Disease in the USA and Europe’, Journal of Veterinary Diagnostic Investigation, 10: 3–10.
Bao, F. et al. (2018) ‘Retrospective Study of Porcine Circovirus Type 2 Infection Reveals a Novel Genotype PCV2f’, Transboundary and Emerging Diseases, 65: 432–40.
Blunt, R. et al. (2011) ‘House Fly Vector for Porcine Circovirus 2b on Commercial Pig Farms’, Veterinary Microbiology, 149: 452–5.
Carman, S. et al. (2006) ‘Porcine Circovirus-2 Associated Disease in Swine in Ontario (2004 to 2005)’, The Canadian Veterinary Journal, 47: 761–2.
Cheung, A. K. et al. (2007) ‘Detection of Two Porcine Circovirus Type 2 Genotypic Groups in United States Swine Herds’, Archives of Virology, 152: 1035–44.
Cortey, M. et al. (2011) ‘Genotypic Shift of Porcine Circovirus Type 2 from PCV-2a to PCV-2b in Spain from 1985 to 2008’, The Veterinary Journal, 187: 363–8.
Davies, B. et al. (2016) ‘Diagnostic Phylogenetics Reveals a New Porcine Circovirus 2 Cluster’, Virus Research, 217: 32–7.
Dupont, K. et al. (2008) ‘Genomic Analysis of PCV2 Isolates from Danish Archives and a Current PMWS Case-Control Study Supports a Shift in Genotypes with Time’, Veterinary Microbiology, 128: 56–64.
Esona, M. et al. (2014) ‘Detection of PCV-2 DNA in Stool Samples from Infants Vaccinated with RotaTeqVR ’, Human Vaccines & Immunotherapeutics, 10: 25–32.
Fenaux, M. et al. (2004a) ‘Two Amino Acid Mutations in the Capsid Protein of Type 2 Porcine Circovirus (PCV2) Enhanced PCV2 Replication In Vitro and Attenuated the Virus In Vivo’, Journal of Virology, 78: 13440–6.
____ et al. (2004b) ‘Detection and In Vitro and In Vivo Characterization of Porcine Circovirus DNA from a Porcine-Derived Commercial Pepsin Product’, The Journal of General Virology, 85: 3377–82.
Franzo, G. et al. (2015) ‘Genetic Characterisation of Porcine Circovirus Type 2 (PCV2) Strains from Feral Pigs in the Brazilian Pantanal: An Opportunity to Reconstruct the History of PCV2 Evolution’, Veterinary Microbiology, 178: 158–62.
____ et al. (2016) ‘Phylodynamic Analysis of Porcine Circovirus Type 2 Reveals Global Waves of Emerging Genotypes and the Circulation of Recombinant Forms’, Molecular Phylogenetics and Evolution, 100: 269–80.
Garcia, L. A. et al. (2012) ‘Surveillance of Human and Swine Adenovirus, Human Norovirus and Swine Circovirus in Water Samples in Santa Catarina, Brazil’, Journal of Water and Health, 10: 445–52.
Gresham, A. et al. (2000) ‘PMWS and Porcine Dermatitis Nephropathy Syndrome in Great Britain’, The Veterinary Record, 146: 143.
Grierson, S. S. et al. (2004) ‘Detection and Genetic Typing of Type 2 Porcine Circoviruses in Archived Pig Tissues from the UK’, Archives of Virology, 149: 1171–83.
Guo, L. J. et al. (2010) ‘Porcine Circovirus Type 2 (PCV2): Genetic Variation and Newly Emerging Genotypes in China’, Virology Journal, 7: 273.
Hamel, A. L., Lin, L. L., and Nayar, G. P. (1998) ‘Nucleotide Sequence of Porcine Circovirus Associated with Postweaning Multisystemic Wasting Syndrome in Pigs’, Journal of Virology, 72: 5262–7.
Harmon, K. et al. (2015) ‘Whole-Genome Sequences of Novel Porcine Circovirus Type 2 Viruses Detected in Swine fromMexico and the United States’, Genome Announcements, 3: e01315–15.
Huang, L. P. et al. (2011) ‘Identification of One Critical Amino Acid That Determines a Conformational Neutralizing Epitope in the Capsid Protein of Porcine Circovirus Type 2’, BMC Microbiology, 11: 188. Jacobsen, B. et al. (2009) ‘Retrospective Study on the Occurrence of Porcine Circovirus 2 Infection and Associated Entities in Northern
Germany’, Veterinary Microbiology, 138: 27–33.
Kim, J., Chung, H. K., and Chae, C. (2003) ‘Association of Porcine Circovirus 2 with Porcine Respiratory Disease Complex’, The Veterinary Journal, 166: 251–6.
Kim, K. H. et al. (2008) ‘Amplification of Uncultured Single-Stranded DNA Viruses from Rice Paddy Soil’, Applied and Environmental Microbiology, 74: 5975–85.
Kiupel, M. et al. (1998) ‘Circovirus-Like Viral Associated Disease in Weaned Pigs in Indiana’, Veterinary Pathology, 35: 303–7.
Kumar, S., Stecher, G., and Tamura, K. (2016) ‘MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets’, Molecular Biology and Evolution, 33: 1870–4.
Kwon, T. et al. (2017) ‘Genotypic Diversity of Porcine Circovirus Type 2 (PCV2) and Genotype Shift to PCV2d in Korean Pig Population’, Virus Research, 228: 24–9.
Labarque, G. G. et al. (2000) ‘Seroprevalence of Porcine Circovirus Types 1 and 2 in the Belgian Pig Population’, The Veterinary Quarterly, 22: 234–6.
Lefebvre, D. J. et al. (2009) ‘Recombination of Two Porcine Circovirus Type 2 Strains’, Archives of Virology, 154: 875–9.
____ et al. (2008) ‘Antigenic Differences among Porcine Circovirus Type 2 Strains, as Demonstrated by the Use of Monoclonal Antibodies’, The Journal of General Virology, 89:
177–87.
Li, L. et al. (2010) ‘Multiple Diverse Circoviruses Infect Farm Animals and Are Commonly Found in Human and Chimpanzee Feces’, Journal of Virology, 84: 1674–82.
Madec, F. et al. (2000) ‘Post-Weaning Multisystemic Wasting Syndrome (PMWS) in Pigs in France: Clinical Observations from Follow-Up Studies on Affected Farms’, Livestock Production Science, 63: 223–33.
Madson, D. M., and Opriessnig, T. (2011) ‘Effect of Porcine Circovirus Type 2 (PCV2) Infection on Reproduction: Disease, Vertical Transmission, Diagnostics and Vaccination’, Animal Health Research Reviews, 12: 47–65.
Magar, R., Mu¨ ller, P., and Larochelle, R. (2000) ‘Retrospective Serological Survey of Antibodies to Porcine Circovirus Type 1 and Type 2’, Canadian Journal of Veterinary Research, 64: 184–6.
Mahe, D. et al. (2000) ‘Differential Recognition of ORF2 Protein from Type 1 and Type 2 Porcine Circoviruses and Identification of Immunorelevant Epitopes’, The Journal of General Virology, 81: 1815–24.
Meerts, P. et al. (2004) ‘Prevalence of Porcine Circovirus 2 (PCV2)-Related Wasting on Belgian Farms with or without a History of Postweaning Multisystemic Wasting Syndrome’, Vlaams Diergeneeskundig Tijdschrift, 73: 31–8.
____ et al. (2005) ‘Replication Kinetics of Different Porcine Circovirus 2 Strains in PK-15 Cells, Fetal Cardiomyocytes and Macrophages’, Archives of Virology, 150: 427–41.
Murrell, B. et al. (2012) ‘Detecting Individual Sites Subject to Episodic Diversifying Selection’, PLoS Genetics, 8: e1002764.
____ et al. (2013) ‘FUBAR: A Fast, Unconstrained Bayesian AppRoximation for Inferring Selection’, Molecular Biology and Evolution, 30: 1196–205.
Nawagitgul, P. et al. (2000) ‘Open Reading Frame 2 of Porcine Circovirus Type 2 Encodes a Major Capsid Protein’, The Journal of General Virology, 81: 2281–7.
Olvera, A., Cortey, M., and Segale´s, J. (2007) ‘Molecular Evolution of Porcine Circovirus Type 2 Genomes: Phylogeny and Clonality’, Virology, 357: 175–85.
Opriessnig, T. et al. (2008) ‘Differences in Virulence among Porcine Circovirus Type 2 Isolates Are Unrelated to Cluster Type 2a or 2b and Prior Infection Provides Heterologous Protection’, The Journal of General Virology, 89: 2482–91.
____, Meng, X. J., andHalbur, P. G. (2007) ‘Porcine Circovirus Type 2 Associated Disease: Update on Current Terminology, Clinical Manifestations, Pathogenesis, Diagnosis, and Intervention Strategies’, Journal of Veterinary Diagnostic Investigation, 19: 591–615.
Pond, S. L., and Frost, S. D. (2005) ‘Datamonkey: Rapid Detection of Selective Pressure on Individual Sites of Codon Alignments’, Bioinformatics, 21: 2531–3.
Ramos, N. et al. (2017) ‘Molecular Study of Porcine Circovirus Type 2 in Wild Boars and Domestic Pigs in Uruguay from 2010 to 2014: Predominance of Recombinant Circulating Strains’, Gene, 637: 230–8.
Reiner, G., Hofmeister, R., and Willems, H. (2015) ‘Genetic Variability of Porcine Circovirus 2 (PCV2) Field Isolates from Vaccinated and Non- Vaccinated Pig Herds in Germany’, Veterinary Microbiology, 180: 41–8.
Rodriguez-Arrioja, G. M. et al. (2003) ‘Retrospective Study on Porcine Circovirus Type 2 Infection in Pigs from 1985 to 1997 in Spain’, Journal of Veterinary Medicine Series B, 50: 99–101.
Rosell, C. et al. (2000) ‘Identification of Porcine Circovirus in Tissues of Pigs with Porcine Dermatitis and Nephropathy Syndrome’, The Veterinary Record, 146: 40–3.
Roy, A., Kucukural, A., and Zhang, Y. (2010) ‘I-TASSER: A Unified Platform for Automated Protein Structure and Function Prediction’, Nature Protocols, 5: 725–38.
Saha, D. et al. (2010) ‘Pathologic and Virologic Findings in Mid-Gestational Porcine Foetuses after Experimental Inoculation with PCV2a or PCV2b’, VeterinaryMicrobiology, 145: 62–8.
____ et al. (2011) ‘Outcome of Experimental Porcine Circovirus Type 1 Infections in Mid- Gestational Porcine Foetuses’, BMC Veterinary Research, 7: 64.
____ et al. (2012a) ‘Single Amino Acid Mutations in the Capsid Switch the Neutralization Phenotype of Porcine Circovirus 2, The ’, Journal of General Virology, 93: 1548–55.
____ et al. (2012b) ‘Antigenic Subtyping and Epitopes’ Competition Analysis of Porcine Circovirus Type 2 Using Monoclonal Antibodies’, Veterinary Microbiology, 157: 13–22. ____ et al. (2014) ‘Unusual Outcome of In Utero Infection and Subsequent Postnatal Super- Infection with Different PCV2b Strains’, Virologica Sinica, 29: 176–82.
Sanchez, R. E., Jr. et al. (2001b) ‘Porcine Circovirus 2 Infection in Swine Foetuses Inoculated at Different Stages of Gestation’, Veterinary Microbiology, 83: 169–76. Replication
in Lymphoid Organs of Pigs Inoculated in Late Gestation or Postnatally and Possible Relation to Clinical and Pathological Outcome of Infection’, Journal of Veterinary Diagnostic Investigation, 16: 175–85.
Sanchez, R., Nauwynck, H., and Pensaert, M. (2001a) ‘Serological Survey of Porcine Circovirus 2 Antibodies in Domestic and Feral Pig Populations in Belgium’, in Proc. ssDNA Viruses of Plants, Birds, Pigs and Primates, St. Malo, France, p. 122.
Schneidman, D. D. et al. (2005) ‘PatchDock and SymmDock: Servers for Rigid and Symmetric Docking’, Nucleic Acids Research, 33: 363–7.
Segales, J., and Domingo, M. (2002) ‘Postweaning Multisystemi Wasting Syndrome (PMWS) in Pigs. A Review’, The Veterinary Quarterly, 24: 109–24.
____ et al. (2008) ‘PCV-2 Genotype Definition and Nomenclature’, The Veterinary Record, 162: 867–8.
____ , Kekarainen, T., and Cortey, M. (2013) ‘The Natural History of Porcine Circovirus Type 2: From an Inoffensive Virus to a Devastating Swine Disease?’, Veterinary Microbiology, 165: 13–20.
Shang, S. B. et al. (2009) ‘Fine Mapping of Antigenic Epitopes on Capsid Proteins of Porcine Circovirus, and Antigenic Phenotype of Porcine
Circovirus Type 2’, Molecular Immunology, 46: 327–34.
Thangthamniyom, N. et al. (2017) ‘Genetic Diversity of Porcine Circovirus Type 2 (PCV2) in Thailand during 2009–2015’, Veterinary Microbiology, 208: 239–46.
Timmusk, S. et al. (2008) ‘Phylogenetic Analysis of Porcine Circovirus Type 2 (PCV2) Pre- and Post-Epizootic Postweaning Multisystemic Wasting Syndrome (PMWS)’, Virus Genes, 36: 509–20.
Trible, B. R. et al. (2011) ‘Antibody Recognition of Porcine Circovirus Type 2 Capsid Protein Epitopes after Vaccination, Infection, and Disease’, Clinical and Vaccine Immunology, 18: 749–57.
____ et al. (2012) ‘Recognition of the Different Structural Forms of the Capsid Protein Determines the Outcome Following Infection with Porcine Circovirus Type 2’, Journal of Virology, 86: 13508-14.
Verreault, D. et al. (2010) ‘Airborne Porcine Circovirus in Canadian Swine Confinement Buildings’, Veterinary Microbiology, 141: 224–30.
Walker, I. W. et al. (2000) ‘Development and Application of a Competitive Enzyme-Linked Immunosorbent Assay for the Detection of Serum Antibodies to Porcine Circovirus Type 2’, Journal of Veterinary Diagnostic Investigation, 12: 400–5.
Wei, C. et al. (2013) ‘Genetic Evolution and Phylogenetic Analysis of Porcine Circovirus Type 2 Infections in Southern China from 2011 to 2012’, Infection, Genetics and Evolution, 17: 87–92.
Wei, R. et al. (2018) ‘Breed Differences in PCV2 Uptake and Disintegration in Porcine Monocytes’, Viruses, 10: 562.
Wiederkehr, D. D. et al. (2009) ‘A New Emerging Genotype Subgroup within PCV-2b Dominates the PMWS Epizooty in Switzerland’, Veterinary Microbiology, 136: 27–35.
Xiao, C. T., Halbur, P. G., and Opriessnig, T. (2015) ‘Global Molecular Genetic Analysis of Porcine Circovirus Type 2 (PCV2) Sequences Confirms the Presence of Four Main PCV2 Genotypes and Reveals a Rapid Increase of PCV2d’, The Journal of General Virology, 96: 1830–41.
Yang, J. et al. (2015) ‘The I-TASSER Suite: Protein Structure and Function Prediction’, Nature Methods, 12: 7–8.
Zhang, Y. (2008) ‘I-TASSER Server for Protein 3D Structure Prediction’, BMC Bioinformatics, 9: 40.
[/registrados]