Desde entonces el virus de la influenza porcina (SIV, de sus siglas en inglés) no solo se ha mantenido en la población porcina, sino que ha aumentado su diversidad.
Sus proteínas externas, Hemaglutinina (H) y Neuraminidasa (N), son las responsables principales de la infección, la patogenicidad y el desarrollo de la respuesta inmunitaria por parte del huésped. Estas proteínas también se emplean para clasificar al virus de la influenza en diferentes subtipos.
En Europa los tres principales subtipos de influenza porcina involucrados en la enfermedad en porcino son el H1N1, el H1N2 y el H3N2.
El SIV -virus de la influenza porcina- ya es de por sí un virus cambiante, porque por un lado es un virus con una cadena simple de ARN que con frecuencia sufre alteraciones al transcribir su cadena de nucleótidos (lo que se denomina deriva genética o “drift”).
El virus de la influenza porcina es un virus cambiante
Por otro lado, su genoma está dividido en ocho segmentos independientes, lo que permite que cuando dos virus distintos infectan al mismo tiempo a una célula diana, los segmentos se intercambien libremente, favoreciendo la aparición de nuevas variantes del virus (lo que se denomina reordenamiento genético o “Shift”).
Aparte del shift y el drift propios del virus de la influenza, la especie porcina tiene una particularidad adicional, porque puede verse afectada por dos de las especies que tienen más importancia en la difusión y variabilidad del virus: la especie humana y las aves.
Esto ocurre porque en el tracto respiratorio de los suínos se expresan receptores para el virus de la influenza procedente tanto de la especie aviar como de la humana.
Los receptores α2-3 se encuentran en aves y equinos, mientras que los α2-6 están en la especie humana y otros mamíferos. Esta configuración de los receptores hace que aparezcan barreras en la transmisión del virus entre especies.
Por ejemplo, la posibilidad de que los équidos o los humanos sean infectados por virus de origen aviar (y viceversa) es muy baja.
Sin embargo, la especie porcina, al expresar los dos tipos de receptores, es susceptible a la infección de virus procedentes tanto de aves como de humanos.
Con el tiempo, las co-infecciones con virus distintos pueden llevar al desarrollo de nuevos virus, a partir de la deriva y el reordenamiento genético.
De esta forma, nos podemos encontrar que una vez adaptado al cerdo, un virus de la influenza originario de otra especie sufra cambios en algunos receptores que le permitan afectar a especies distintas.
Por ejemplo, un SIV originariamente aviar puede acabar siendo capaz de infectar a la especie humana si emplea al cerdo como huésped intermedio.
Por lo tanto, podemos decir que el cerdo puede sufrir la infección del virus de distintas especies y ejercer de “efecto coctelera”, siendo potencialmente el puente de la transmisión de SIV entre aves y humanos.

Sin embargo, en continentes como Europa y América, donde la producción está más industrializada y especializada, esta fuente de variación no es tan alta. Además, debido a la segregación entre ambos continentes, el SIV ha evolucionado de forma distinta en Europa y América.


- A finales de los 70, fueron las aves las que introdujeron una nueva variante -un H1N1 enteramente de origen aviar que reemplazó al H1huN1 original-
- Más tarde, a mediados de los 80, empezó a circular el subtipo H3N2 de origen humano, proveniente de la reordenación de un H3N2 humano (de Asia) y un H1N1 aviar, con genes porcinos adaptados
- A mediados de los 90 apareció el subtipo H1huN2, que en este caso proviene de una triple reordenación (shift), entre genes aviares, humanos y porcinos
APARICIÓN DEL VIRUS H1N1 PANDÉMICO (H1PANN1)
El H1panN1 que generó tanta alarma a nivel mundial en 2009, es el producto de diversas reordenaciones genéticas entre SIV de origen aviar, humano y porcino que sucedieron en la cabaña porcina norteamericana durante más de diez años.
En América no se llegó a introducir el H1avN1, por lo que el subtipo H1N1 mayoritario continuaba siendo un H1N1 clásico (H1huN1 o H1csN1). El H3N2 fue detectado en los 70, pero no se consideró un virus demasiado patógeno.
A finales de la década de los 90 se sucedieron una serie de reordenaciones que aumentaron la diversidad del SIV en Norteamérica:
- En un primer paso se generó un H3N2 triple recombinante (trH3N2) a partir de la reordenación de un H1csN1, un virus aviar y un H3N2 humano de la gripe estacional.
- En un segundo paso, este nuevo trH3N2 se reordenó otra vez con un H1csN1, adaptando su HA (resultando en un trH1N2) o su HA y NA (resultando en un trH1N1).
- Finalmente, cerca de 2009, el trH1N2 se reordenó con un H1avN1 de origen euroasiático, que aportó los genes de la NA y la matriz, apareciendo el H1panN1.
Los ocho segmentos del H1panN1 son similares a virus porcinos. Esto, junto con la capacidad del porcino para recibir la infección del virus de origen aviar y humano, ha llevado a asumir que este nuevo virus ha emergido del cerdo.
La distribución mundial del H1panN1 la ha llevado a cabo la especie humana y de forma retrógrada la ha acabado introduciendo en las explotaciones porcinas. Además, el H1panN1 es ahora fuente de nuevas reordenaciones, como el H1panN2 que ya se está diagnosticando en varios países europeos.
Estudios realizados en el CReSA en 2011 en España, demuestran que la prevalencia de granjas positivas a anticuerpos (han estado en contacto con el virus) es muy alta.
El 94% de las explotaciones son positivas al menos a un subtipo y más del 40% son positivas como mínimo a dos subtipos distintos. Más del 10% son positivas a los tres. El subtipo más prevalente fue el H1N1, seguido del H1N2 y el H3N2.
En España la prevalencia de granjas positivas a SIV es muy alta
Sin embargo, actualmente es posible que la situación esté cambiando y que en algunas explotaciones ya se haya introducido el H1panN1. Incluso estamos observando la aparición de nuevas variantes, fruto de la reordenación, como es el caso del H1avN2 (un subtipo con una elevada prevalencia en Dinamarca) que es fruto de la reordenación entre H1avN1 y H1huN2 o H3huN2.
Otro ejemplo sería el H3huN1 detectado en Alemania, que sería fruto de la reordenación entre H1avN1 y H3huN2.


Estamos viendo que el SIV está en evolución constante, lo que implica que tiene una elevada variabilidad antigénica, con cepas de distinta similitud incluso dentro de un mismo subtipo.
La protección cruzada entre distintos subtipos suele ser baja y en ocasiones incluso entre las distintas cepas de un mismo subtipo puede ser parcial.
En el caso de la H1, los tres tipos de H1 presentes hoy en día (H1avN1, H1huN2, y H1N1pdm) a nivel antigénico prácticamente no muestran reacciones cruzadas porque provienen de orígenes distintos (aviar, humano y porcino en Norteamérica).
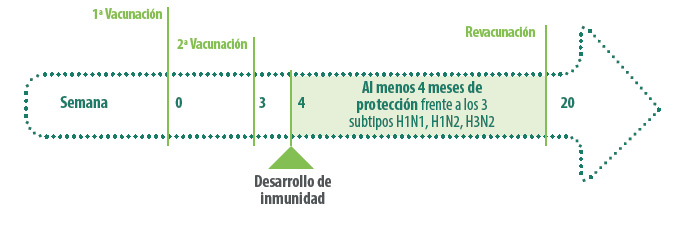
Esto obliga a que las vacunas tengan que ponerse al día periódicamente, introduciendo los nuevos subtipos circulantes
Tal como hemos explicado, una vacuna que contenga los tres principales subtipos H1avN1, H1huN2 y H3N2 no protegerá completamente contra un H1panN1, pero sí que lo hará contra reordenaciones entre los tres subtipos anteriores, como serían el H1avN2 o el H3huN1, ya que la vacuna ofrece buena protección contra sus HA y NA de forma independiente.
Otro aspecto a tener en cuenta es la necesidad de actualizar las cepas presentes en las vacunas. Sabemos que en la especie humana, las vacunas se están actualizando anualmente frente a las cepas de gripe circulantes.
Sin embargo, esto no es factible en el caso del porcino en Europa, porque el proceso de registro de una vacuna requiere un mínimo de entre cinco y diez años.
Evidentemente, el grado de protección es mejor cuanto más se parece la cepa vacunal a la cepa de campo, pero hay que tener en cuenta que la variabilidad del virus influenza humano es mayor debido al mayor tamaño y movilidad de la población humana mundial, lo que da lugar a una mayor deriva genética.
En porcino, el virus de la influenza no se ve obligado a mutar tanto, ya que encuentra de manera fácil animales susceptibles
La deriva genética en el cerdo parece menor que en humanos, probablemente en parte debido al flujo constante de animales susceptibles a la infección que representa el nacimiento constante de lechones y la introducción de reemplazos.
En esta situación, el SIV no se ve obligado a mutar tanto porque encuentra fácilmente animales susceptibles a la infección.
Si se dieran mutaciones que ofrecieran una ventaja (por ejemplo escaparan mejor a la respuesta inmune), estas no se establecerían con tanta facilidad como en la especie humana porque no hay tanta necesidad para ello.
Sin embargo, tal como hemos visto en este artículo, las reordenaciones sí que parece que suceden de forma habitual en la cabaña porcina, a diferencia de los virus humanos, que en mayor medida evolucionan gracias a la deriva, lo que obliga a actualizar con más frecuencia las vacunas detinadas a la especie humana.
Otra diferencia de la vacunación en la especie humana frente a la porcina es la capacidad de establecer mejores programas vacunales.
Mientras que las campañas vacunales en humanos recomiendan la vacunación de los jóvenes y ancianos una vez al año (y es probable que una buena parte de los individuos de estos grupos de riesgo no se vacunen…), en una explotación porcina podemos decidir con qué frecuencia queremos vacunar.
Y si queremos, podemos vacunar a todos los animales al mismo tiempo. Esto genera un grado de protección uniforme que contribuye a dificultar la circulación del virus, reducir las subpoblaciones y mantener la enfermedad bajo control.
El SIV es un virus muy difundido, dinámico, con una alta diversidad genética y antigénica y que está en evolución constante.
En los últimos 40 años se han introducido cuatro variantes distintas en la población porcina europea (el H1avN1, el H1huN1, el H3huN2 y el H1panN1), lo que significa que aparece una nueva cada diez años como media.
Esto obliga a actualizar las vacunas comerciales periódicamente para poder garantizar una buena eficacia contra la enfermedad