

El envejecimiento ovárico en las cerdas causa infertilidad y trastornos reproductivos. Factores como el estrés oxidativo, la ferroptosis y la autofagia parecen estar implicados en este proceso, pero los mecanismos subyacentes no se comprenden del todo. Este estudio se ha centrado en identificar los cambios moleculares y genéticos relacionados con el envejecimiento ovárico en cerdas, con el objetivo de aportar conocimiento para el desarrollo de estrategias que ayuden a mitigar sus efectos.
ENVEJECIMIENTO OVÁRICO Y ESTRÉS OXIDATIVO
El envejecimiento ovárico se caracteriza por la reducción del número de folículos y un deterioro de la calidad de los ovocitos, lo que conduce a una disminución gradual de la fertilidad de las hembras1,2.
ESTRÉS OXIDATIVO
En los mamíferos, las causas del envejecimiento ovárico se atribuyen principalmente al desequilibrio de los sistemas antioxidantes3. El estrés oxidativo es el principal impulsor del envejecimiento ovárico:
![]()

![]()
La acumulación excesiva de especies reactivas del oxígeno (ROS) inducida por el estrés oxidativo provoca la apoptosis de las células de la granulosa (CG)6, potencia la degeneración del cuerpo lúteo y conduce a la atresia folicular en las cerdas7.
FERROPTOSIS
La ferroptosis es una forma de muerte celular no apoptótica dependiente del hierro (Fe) y de las ROS8 caracterizada por la acumulación de especies reactivas de oxígeno lipídicas y hierro, formando así hidroperóxidos lipídicos9,10.
AUTOFAGIA
La autofagia, uno de los principales tipos de muerte celular programada11 y muerte celular de tipo II, consiste en la digestión las proteínas mal plegadas en los lisosomas, limpiando los orgánulos dañados, como las mitocondrias y los peroxisomas12.
![]()
![]()
Las observaciones realizadas en diversos estudios17-22 con otros diversos tipos celulares sugieren que el estrés oxidativo podría inducir la ferroptosis y la autofagia para regular el envejecimiento ovárico, pero los mecanismos implicados aún no se han dilucidado.
FOSFOLÍPIDO FOSFATASA 3
La fosfolípido fosfatasa 3 (PLPP3) pertenece a la familia de las fosfatasas del ácido fosfatídico y desfosforila los fosfolípidos en productos de señalización lipídica que suelen ser oxidadas por las ROS dando lugar a hidroperóxidos lipídicos23.
Estudios previos han revelado que los hidroperóxidos lipídicos inducen apoptosis celular, ferroptosis y autofagia17,24.
Por tanto, la PLPP3 podría estar implicada en el envejecimiento ovárico a través de la regulación del estrés oxidativo, la ferroptosis y la autofagia.

PAPEL DE PLPP3 EN EL ENVEJECIMIENTO OVÁRICO
En este estudio, se trató de explorar las funciones biológicas de PLPP3 en el estrés oxidativo, la ferroptosis y la autofagia en las CG e investigar el papel de PLPP3 en el desarrollo folicular.
Dado que el inicio de la pubertad indica la maduración de los folículos porcinos, se identificaron además las asociaciones entre la edad al inicio de la pubertad y los posibles marcadores moleculares de PLPP3. Estos resultados podrían proporcionar nuevas dianas para las investigaciones sobre el envejecimiento ovárico en mamíferos.
MATERIALES Y MÉTODOS
OBTENCIÓN Y ANÁLISIS DE LAS CG
![]()

Para este estudio se utilizaron ovarios envejecidos obtenidos de cerdas sacrificadas (más de 6 partos) y ovarios jóvenes obtenidos de cerdas de 5 meses.
Las células de la granulosa (CG) primarias porcinas se cultivaron para explorar el impacto de la PLPP3 en sus funciones celulares.
![]()

DETECCIÓN DE LA PUBERTAD Y ANÁLISIS GENÉTICO
![]()
| El inicio de la pubertad de las cerdas se identificó mediante la observación del reflejo de inmovilidad y por contacto con los verracos, registrándose los signos de pubertad dos veces al día.
Se extrajo ADN de 142 cerdas para calcular la frecuencia de genotipos y la frecuencia alélica de los SNPs en el promotor de PLPP3. |
Seguidamente, se realizó el análisis de asociación genética entre los SNPs PLPP3 y de la edad de presentación de la pubertad de las cerdas. |
DETECCIÓN ROS, AUTOFAGIA, SA β-GAL, MDA Y FE
![]()
Con el fin de caracterizar el estrés oxidativo y envejecimiento celular de las CG se realizaron los siguientes análisis:
- Detección de ROS.
- Detección del nivel de autofagia.
- Prueba SA β-Gal.
- Detección de malondialdehído (MDA).
- Detección de hierro (Fe)
- Determinación del potencial de membrana mitocondrial
- Ensayo Western Blot (WB)
RESULTADOS
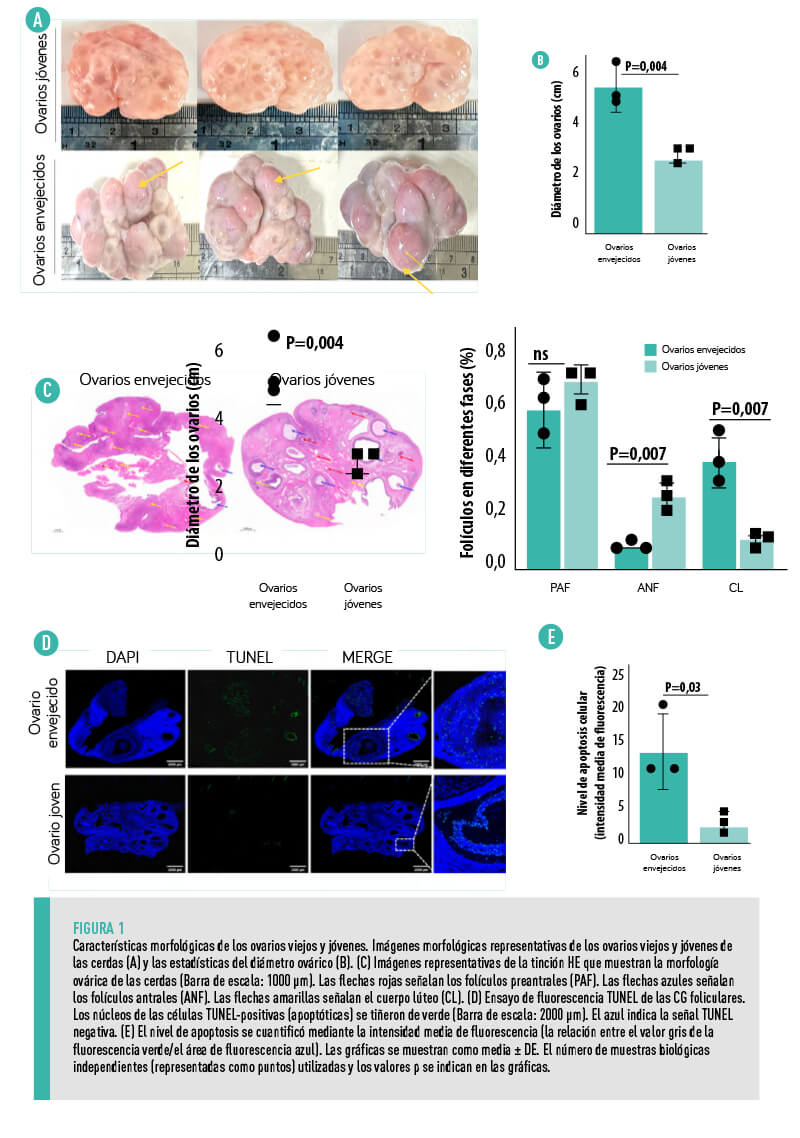
CARACTERÍSTICAS MORFOLÓGICAS DE LOS OVARIOS
![]()
El tamaño [registrados]de los ovarios envejecidos (4-6 cm de diámetro) era mayor que el de los jóvenes (2-3 cm de diámetro) (Figura 1A,B), presentando los primeros mayor abundancia de cuerpos lúteos (flechas amarillas) (Figura 1A). Además, en comparación con los ovarios envejecidos, los ovarios jóvenes tenían más folículos primarios (flechas rojas) y antrales (flechas azules), con un aumento significativo de los folículos antrales (Figura 1C).
Cabe destacar que los ovarios envejecidos también presentaban mayores niveles de apoptosis de CG en los folículos antrales que los jóvenes (Figura 1D,E).

| Los niveles de ARNm de la superóxido dismutasa 1 y 2 (SOD1, SOD2), la proteína 1 beta de cadena ligera asociada a microtúbulos (LC3B) y los marcadores asociados al envejecimiento sirtuina 1 (SIRT1) y sirtuina 6 (SIRT6) se encontraron notablemente reducidos en los ovarios envejecidos (Figura 2A).
Los niveles de ARNm del citocromo P450 familia 1 subfamilia B miembro 1 (CYP1B1), relacionado con la autofagia 7 (ATG7), relacionado con la autofagia 5 (ATG5), beclina 1 (BECN1), SQSTM1 sequestosoma 1 (p62), y el inhibidor de la quinasa dependiente de ciclina 2A (p16) estuvieron significativamente aumentados (Figura 2A). Los niveles proteicos de SOD1, p62, SIRT1 y SIRT6 disminuyeron notablemente en los ovarios envejecidos, pero los de p62 y ATG7 aumentaron en comparación con los de los ovarios jóvenes (Figura 2B,C). |
ESTOS RESULTADOS SUGIEREN QUE EL ESTRÉS OXIDATIVO Y LA AUTOFAGIA PODRÍAN REGULAR EL ENVEJECIMIENTO OVÁRICO |
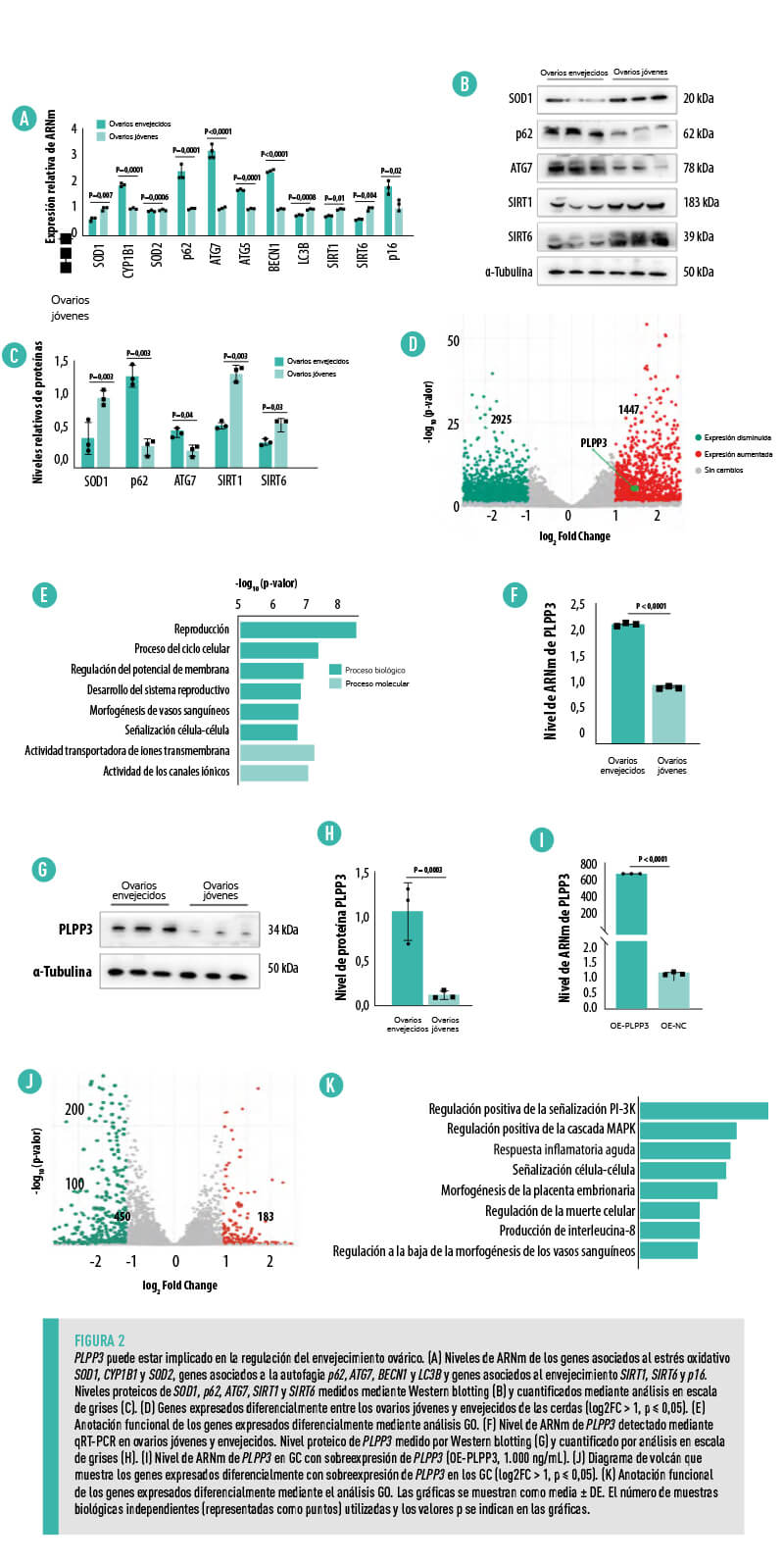

La secuenciación de ARN reveló que 1.447 genes tenían una expresión significativamente aumentada y 2.925 genes tenían expresión disminuida en los ovarios envejecidos en comparación con los jóvenes (Figura 2D).
El análisis de Ontología Génica (GO) reveló que los genes expresados diferencialmente estaban enriquecidos principalmente en la regulación del proceso reproductivo y el potencial de membrana, habiendo sugerido estudios previos que la disminución del potencial de membrana provocada por la disfunción mitocondrial contribuye a la senescencia celular25 (Figura 2E).
LA EXPRESIÓN DE PLPP3 ESTABA SIGNIFICATIVAMENTE AUMENTADA EN LOS OVARIOS ENVEJECIDOS EN COMPARACIÓN CON LOS JÓVENES (FIGURA 2D)
A diferencia de los ovarios jóvenes, los niveles de ARNm (Figura 2F) y proteína (Figura 2G,H) de PLPP3 aumentaron notablemente en los ovarios envejecidos y la sobreexpresión de PLPP3 (Figura 2I) aumentó la expresión de 183 genes y redujo la de 450 genes en las CG en comparación con el grupo de control (Figura 2J).
| Estos genes expresados de forma diferencial estaban principalmente enriquecidos en la regulación de la respuesta inflamatoria, la morfogénesis de los vasos sanguíneos, la señalización MAPK y PI- 3K (Figura 2K), esenciales en la regulación del desarrollo folicular y el envejecimiento ovárico26,27. |
PLPP3 AUMENTA LA SENESCENCIA DE LAS CG
![]()
Se utilizó la sobreexpresión de PLPP3 (Figura 2I) y el knockdown (Figura 3A) para explorar su impacto en la senescencia celular de las CG primarias porcinas.
- La sobreexpresión de PLPP3 disminuyó significativamente el potencial de membrana mitocondrial en las GC (Figura 3B,C).
- La sobreexpresión de PLPP3 aumentó la actividad SA-β-Gal en las CG, mientras que las CG con knockdown de PLPP3 inhibieron eficazmente la actividad SA-β-Gal (Figura 3D,E).
- Los niveles de ARNm (Figura 3F) y proteína (Figura 3G,H) de los marcadores asociados a la senescencia SIRT1 y SIRT6 estaban significativamente suprimidos en las CG con sobreexpresión de PLPP3. Al mismo tiempo, aumentó la expresión del inhibidor de la cinasa dependiente de ciclina 1A (p21).
| En cambio, el knockdown de PLPP3 estimuló notablemente la expresión de SIRT1 y SIRT6 e inhibió p21. |
- Los resultados de la inmunofluorescencia mostraron que la proteína PLPP3 estaba aumentada en los ovarios envejecidos en comparación con los ovarios jóvenes, mientras que SIRT1 estaba regulada a la baja (Figura 3I).
EN GENERAL, ESTOS RESULTADOS INDICAN QUE PLPP3 PODRÍA AUMENTAR LA SENESCENCIA CELULAR EN LAS CG
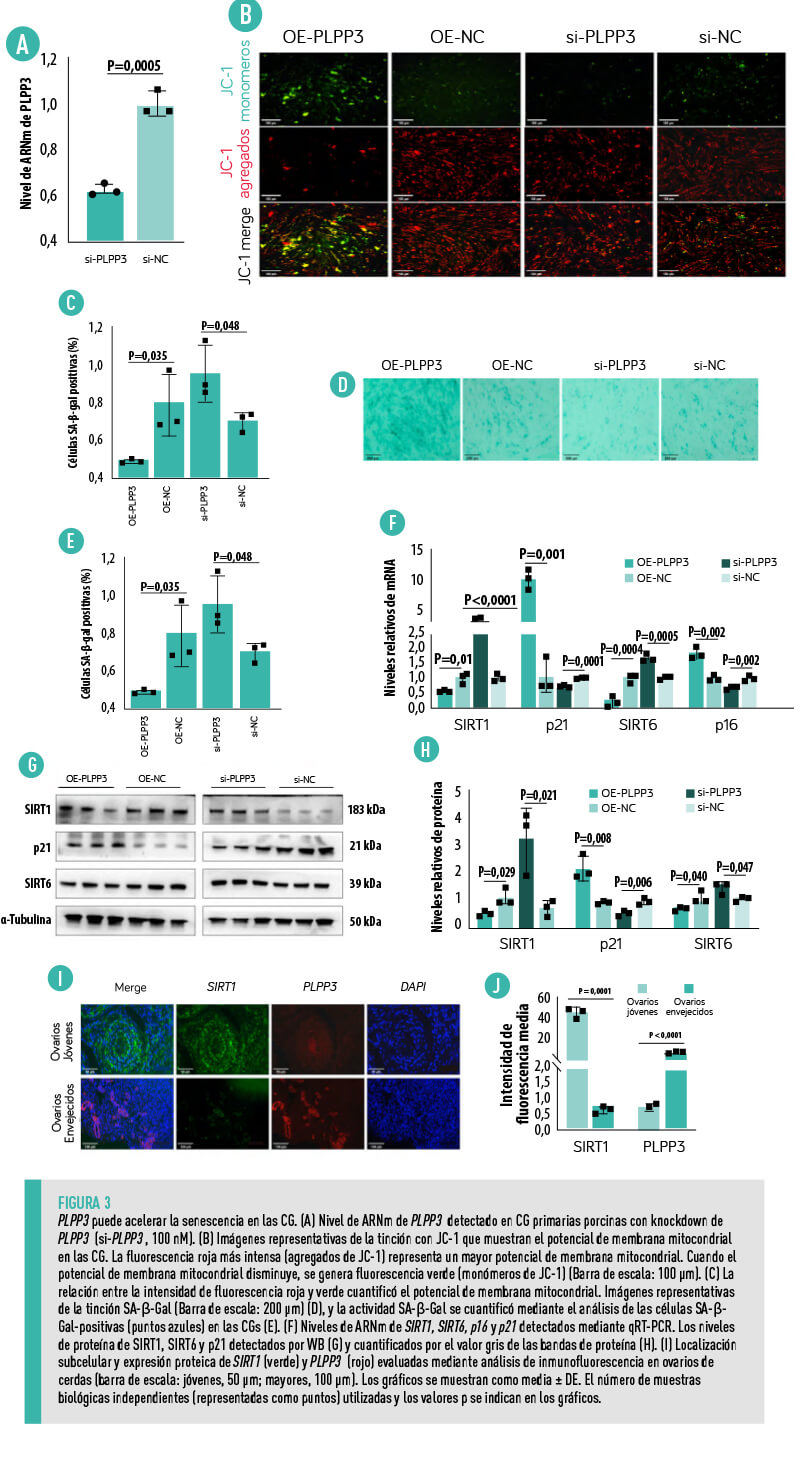

PLPP3 AGRAVA EL ESTRÉS OXIDATIVO Y LA FERROPTOSIS DE LAS CG
![]()
- La sobreexpresión de PLPP3 aumentó significativamente la acumulación de ROS intracelulares en las CG (Figura 4A,B), mientras que el knockdown de PLPP3 disminuyó notablemente los niveles de ROS.
- Los niveles de ARNm de SOD1, CAT y SOD2 se redujeron significativamente en las CG con sobreexpresión de PLPP3, acompañados de un aumento de CYP1B1 (Figura 4C).
- La sobreexpresión de PLPP3 redujo notablemente los niveles proteicos de SOD1 y CAT, mientras que se observó un aumento de la expresión de SOD1 y CAT en las CG con knockdown de PLPP3 (Figura 4D,E).
- Los resultados de la inmunofluorescencia revelaron que la expresión de PLPP3 estaba aumentada en los ovarios envejecidos de las cerdas, pero acompañada de disminución de la expresión de SOD1 (Figura 4F).
- El malondialdehído (MDA), un marcador de la peroxidación lipídica inducida por el estrés oxidativo presentó unos niveles significativamente mayores las CG con sobreexpresión de PLPP3 (Figura 4G).
- La sobreexpresión de PLPP3 aumentó el nivel de Fe (Figura 4H), lo que puede promover la peroxidación lipídica y la acumulación de ROS y disminuir el potencial de membrana mitocondrial28.
| En cambio, el knockdown de PLPP3 disminuyó significativamente, tanto los niveles de MDA como de Fe en las CG (Figura 4G,H). |
- Los niveles de ARNm de los genes supresores de la ferroptosis glutatión peroxidasa 4 (GPX4), ferritina cadena pesada 1 (FTH1) y proteína supresora de la ferroptosis 1 (FSP1) se redujeron notablemente en las CG con sobreexpresión de PLPP3, pero el nivel de ARNm de la NADPH oxidasa 1 (NOX1) aumentó (Figura 4I).
- La sobreexpresión de PLPP3 redujo significativamente los niveles de las proteínas supresoras de la ferroptosis FSP1, FTH1 y del miembro 11 de la familia 7 de transportadores de solutos (SLC7A11), mientras que el knockdown de PLPP3 aumentó de forma evidente la expresión de FSP1, FTH1 y SLC7A11 (Figura 4J,K).
ESTOS RESULTADOS SUGIEREN QUE PLPP3 PODRÍA PROVOCAR ESTRÉS OXIDATIVO Y FERROPTOSIS PARA ACELERAR LA SENESCENCIA EN LAS CG
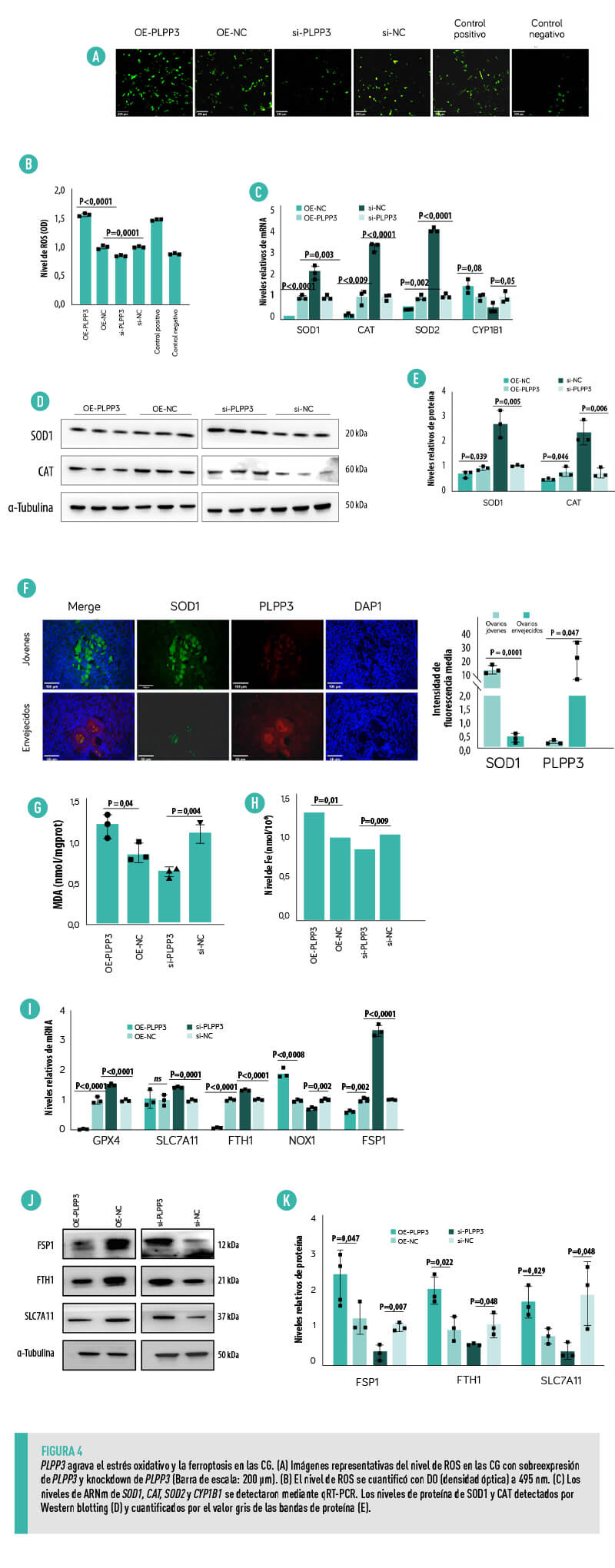

PLPP3 ELEVA EL NIVEL DE AUTOFAGIA EN L AS CG
![]()
Se empleó el adenovirus mRFP-GFP-LC3 en tándem para medir el impacto de PLPP3 en la autofagia de las CG.
- La sobreexpresión de PLPP3 incrementó notablemente el número de autofagosomas y autolisosomas, aumentando la tasa de autofagia (Figura 5A,B).
| En cambio, el knockdown de PLPP3 atenuó significativamente dicha autofagia al reducir el número de autofagosomas y autolisosomas (Figura 5A,B). |
- La sobreexpresión de PLPP3 aumentó los niveles de ARNm de LC3B, ATG7, BECN1 y ATG5, mientras que disminuyó el nivel de ARNm de p62 (Figura 5C).
- Los niveles de proteína de LC3B y ATG7 aumentaron notablemente con la sobreexpresión de PLPP3, mientras que el nivel de proteína de p62 disminuyó notablemente (Figura 5D,E).
- El knockdown de PLPP3 suprimió la expresión de LC3B y ATG7 y estimuló la expresión de p62 (Figura 5D,E), lo que sugiere que la sobreexpresión de PLPP3 podría despertar una respuesta de autofagia protectora frente al daño por estrés oxidativo en las CG.
- Los resultados de la inmunofluorescencia demostraron además que PLPP3 estaba regulada al alza en los ovarios envejecidos y acompañada de una regulación al alza de p62 (Figura 5F), lo que indicaba un bajo nivel de autofagia en los ovarios envejecidos.
ESTOS RESULTADOS SUGIEREN QUE PLPP3 PODRÍA DESENCADENAR LA AUTOFAGIA INDUCIDA POR EL ESTRÉS OXIDATIVO, CONTRIBUYENDO ASÍ AL ENVEJECIMIENTO OVÁRICO DE LAS CERDAS

g.155798586T>C Y g.155798718C>T PODRÍAN SER MARCADORES MOLECULARES DEL DESARROLLO FOLICULAR
![]()
Dado que el inicio de la pubertad es un signo distintivo de la maduración de los folículos29,30, se realizó un análisis de asociación genética entre los SNPs del promotor de PLPP3 y la edad de la pubertad.
El análisis del polimorfismo genético (n = 142) reveló cuatro SNPs en el promotor de PLPP3:
|
|

DISCUSIÓN
En este estudio, se observó que los ovarios envejecidos tenían relativamente más cuerpos lúteos que los jóvenes, mientras que los ovarios jóvenes tenían abundantes folículos primarios y folículos antrales. En los ovarios envejecidos también se observó una disminución del antioxidante SOD1 responsable de la defensa frente al estrés oxidativo inducido por las ROS31.


Se observó que PLPP3 tenía una expresión significativamente más alta en los ovarios envejecidos que en los ovarios jóvenes y revelándose posibles vías a través de las cuales influye en la regulación del desarrollo folicular, como la inflamación, la señalización MAPK y la regulación de la morfogénesis de los vasos sanguíneos.
PLPP3 PODRÍA SER UN POTENCIAL REGULADOR DEL ENVEJECIMIENTO OVÁRICO A TRAVÉS DE LA INDUCCIÓN DEL ESTRÉS OXIDATIVO
El análisis de polimorfismo genético identificó los SNPs g.155798586 T>C y g.155798718 C>T en el promotor de PLPP3 que podrían servir como marcadores moleculares del desarrollo folicular, al mostrar una correlación significativa con la edad de presentación de la pubertad.
![]()

Por otro lado, se demostró que la sobreexpresión de PLPP3 aumenta significativamente la senescencia celular en las CG, junto con un elevado nivel de ROS y una disminución de la expresión de los antioxidantes SOD1 y CAT. Esto es relevante porque las enzimas SOD desempeñan un papel crucial en la eliminación de ROS y son comunes en los ovarios de mamíferos35.
La sobreexpresión de PLPP3 indujo la formación de hidroperóxidos lipídicos y ferroptosis al favorecer la acumulación de MDA y Fe en las CG.
| Dado que el exceso de ROS contribuye en gran medida a la ferroptosis, se sugiere que PLPP3 puede acelerar la senescencia de las CG al alterar el equilibrio entre antioxidantes y ROS para agravar el estrés oxidativo. |
La acumulación de autofagosomas conduce a la muerte celular en las CG foliculares de rata36, mientras que el estrés oxidativo induce respuestas de autofagia, apoptosis e inflamación en mamíferos37,38.
| Si bien, la autofagia moderada es beneficiosa para la supervivencia celular al eliminar los productos oxidados y las mitocondrias dañadas9, un exceso es responsable de la muerte de las CG inducida por estrés oxidativo15. |
En este caso, se encontró que PLPP3 aumentó el nivel de autofagia en las CG, lo que indicaría una exacerbación del estrés oxidativo por PLPP3 como inductor de la autofagia y del envejecimiento ovárico.
| En conclusión, este estudio revela que PLPP3 podría ser un potencial regulador del envejecimiento ovárico. Concretamente, PLPP3 agravó el estrés oxidativo, la ferroptosis y la autofagia en las CG, conduciendo a una aceleración de la senescencia celular.
Además, los SNPs g.155798586 T>C y g.155798718 C>T del promotor de PLPP3 podrían ser marcadores moleculares del desarrollo folicular porcino. Estos resultados pueden aportar información valiosa para investigar los mecanismos del envejecimiento ovárico en mamíferos. |
BIBLIOGRAFÍA
1. Krysko, D.V.; Diez-Fraile, A.; Criel, G.; Svistunov, A.A.; Vandenabeele, P.; D’Herde, K. Life and death of female gametes during oogenesis and folliculogenesis. Apoptosis 2008, 9, 1065–1087. [CrossRef] [PubMed]
2. Broekmans, F.J.; Soules, M.R.; Fauser, B.C. Ovarian aging: Mechanisms and clinical consequences. Endocr. Rev. 2009, 5, 465–493. [CrossRef] [PubMed]
3. Zhang, J.; Chen, Q.; Du, D.;Wu, T.;Wen, J.;Wu, M.; Zhang, Y.; Yan,W.; Zhou, S.; Li, Y.; et al. Can ovarian aging be delayed by pharmacological strategies. Aging 2019, 11, 16. [CrossRef]
4. Yang, L.; Chen, Y.; Liu, Y.; Xing, Y.; Miao, C.; Zhao, Y.; Chang, X.; Zhang, Q. The Role of Oxidative Stress and Natural Antioxidants in Ovarian Aging. Front. Pharmacol. 2020, 11, 617843. [CrossRef]
5. Wang, L.; Tang, J.; Wang, L.; Tan, F.; Song, H.; Zhou, J.; Li, F. Oxidative stress in oocyte aging and female reproduction. J. Cell Physiol. 2021, 236, 7966–7983. [CrossRef]
6. Tiwari, M.; Prasad, S.; Tripathi, A.; Pandey, A.N.; Ali, I.; Singh, A.K.; Shrivastav, T.G.; Chaube, S.K. Apoptosis in mammalian oocytes: A review. Apoptosis 2015, 20, 1019–1025. [CrossRef] [PubMed]
7. Meng, L.; Wu, Z.; Zhao, K.; Tao, J.; Chit, T.; Zhang, S.; Wang, C.C.; Teerds, K. Transcriptome Analysis of Porcine Granulosa Cells in Healthy and Atretic Follicles: Role of Steroidogenesis and Oxidative Stress. Antioxidants 2020, 10, 22. [CrossRef]
8. Shenghui Zhang, Q.L.; Chang, M.; Pan, Y.; Yahaya, B.H.; Liu, Y.; Lin, J. Chemotherapy impairs ovarian function through excessive ROS-induced ferroptosis. Cell Death Dis. 2022, 14, 340. [CrossRef]
9. Scott, J.; Dixon, K.M.L.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An irondependent form of nonapoptotic cell death. Cell 2022, 149, 1060. [CrossRef]
10. Li, J.; Cao, F.; Yin, H.-L.; Huang, Z.-J.; Lin, Z.-T.; Mao, N.; Sun, B.;Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [CrossRef] [PubMed]
11. Baehrecke, E.H. Autophagy: Dual roles in life and death? Nat. Rev. Mol. Cell Biol. 2005, 6, 505–510. [CrossRef] [PubMed]
12. Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [CrossRef]
13. Yahyah Ama, T.S.-M.; Hansen, M.; Morimoto, R.I.; Simon, A.K.; Bjedov, I.; Tavernarakis, N.; Rubinsztein, D.C.; Partridge, L.; Kroemer, G.; Labbadia, J.; et al. Autophagy in healthy aging and disease. Nat. Aging 2021, 1, 17. [CrossRef]
14. Andrew, M.; Leidal, B.L.; Debnath, J. Autophagy and the cell biology of age-related disease. Nat. Cell Biol. 2018, 20, 11. [CrossRef]
15. Zhang, J.Q.; Ren, Q.L.; Chen, J.F.; Gao, B.W.; Wang, X.W.; Zhang, Z.J.; Wang, J.; Xu, Z.J.; Xing, B.S. Autophagy Contributes to Oxidative Stress-Induced Apoptosis in Porcine Granulosa Cells. Reprod. Sci. 2020, 28, 14. [CrossRef]
16. Lu, J.; Huang, J.; Zhao, S.; Xu,W.; Chen, Y.; Li, Y.;Wang, Z.; Dong, Y.; You, R.; Cao, J.; et al. FOXO1 Is a Critical Switch Molecule for Autophagy and Apoptosis of Sow Endometrial Epithelial Cells Caused by Oxidative Stress. Oxid. Med. Cell Longev. 2021, 2021, 1172273. [CrossRef]
17. Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell Longev. 2019, 2019, 5080843. [CrossRef] [PubMed]
18. Huiqin Zhong, M.X.; Zarkovic, K.; Zhu, M.; Sa, R.; Lu, J.; Tao, Y. Mitochondrial control of apoptosis through modulation of cardiolipin oxidation in hepatocellular carcinoma_ A novel link between oxidative stress and cancer. Free Radic. Biol. Med. 2017, 102, 10. [CrossRef]
19. Vernon, W.; Dolinsky, A.Y.M.C.; Frayne, I.R.; Light, P.E.; Rosiers, C.D.; Dyck, J.R.B. Resveratrol prevents the prohypertrophic effects of oxidative stress on LKB1. Circ. Res. 2009, 119, 10. [CrossRef]
20. Park, E.; Chung, S.W. ROS-mediated autophagy increases intracellular iron levels and ferroptosis by ferritin and transferrin receptor regulation. Cell Death Dis. 2019, 10, 822. [CrossRef]
21. Gao, M.; Monian, P.; Pan, Q.; Zhang, W.; Xiang, J.; Jiang, X. Ferroptosis is an autophagic cell death process. Cell Res. 2016, 26, 12. [CrossRef [PubMed]
22. Wen Hou, Y.X.; Song, X.; Sun, X.; Lotze, M.T. Autophagy promotes ferroptosis by degradation of ferritin. Autophagy 2016, 12, 7366. [CrossRef]
23. Porter, N.A. Mechanisms for the autoxidation of polyunsaturated lipids. Acc. Chem. Res. 1986, 19, 262–268. [CrossRef]
24. Chen, X.; Li, J.; Kang, R.; Klionsky, D.J.; Tang, D. Ferroptosis: Machinery and regulation. Autophagy 2021, 17, 2054–2081. [CrossRef]
25. Miwa, S.; Kashyap, S.; Chini, E.; von Zglinicki, T. Mitochondrial dysfunction in cell senescence and aging. J. Clin. Investig. 2022, 132, 158447. [CrossRef]
26. Carolina Lliberos, S.H.L.; Zareie, P.; La Gruta, N.L.; Mansell, A.; Hutt, K. Evaluation of inflammation and follicle depletion during ovarian ageing in mice. Sci. Rep. 2021, 11, 278. [CrossRef]
27. Huang, H.M.; Huang, C.J.; Yen, J.J.Y. Mcl-1 is a common target of stem cell factor and interleukin-5 for apoptosis prevention activity via MEK/MAPK and PI-3K/Akt pathways. Blood 2022, 96, 8.
28. Guang-Hui Chen, C.-C.S.; Pantopoulos, K.;Wei, X.-L.; Zheng, H.; Luo, Z. Mitochondrial oxidative stress mediated Fe-induced ferroptosis via the NRF2-ARE pathway. Free Radic. Biol. Med. 2022, 180, 13. [CrossRef]
29. Likszo, P.; Gromadzka-Hliwa, K.; Klos, J.; Kaczmarek, M.M.; Ziecik, A.J. Attainment of Sexual Maturity and Gonadotropin Priming in Gilts Determine Follicular Development, Endocrine Milieu and Response to Ovulatory Triggers. Int. J. Mol. Sci. 2022, 23, 9190. [CrossRef]
30. Pankhurst, M.W. A putative role for anti-Müllerian hormone (AMH) in optimising ovarian reserve expenditure. Eur. J. Endocrinol. 2017, 148, 9. [CrossRef]
31. Racchi, M.L. Antioxidant Defenses in Plants with Attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [CrossRef]
32. Sezer, Z.; Ekiz Yilmaz, T.; Gungor, Z.B.; Kalay, F.; Guzel, E. Effects of vitamin E on nicotine-induced lipid peroxidation in rat granulosa cells: Folliculogenesis. Reprod. Biol. 2020, 20, 63–74. [CrossRef]
33. Pasqualotto, E.B.; Agarwal, A.; Sharma, R.K.; Izzo, V.M.; Pinotti, J.A.; Joshi, N.J.; Rose, B.I. Effect of oxidative stress in follicular fluid on the outcome of assisted reproductive procedures. Fertil. Steril. 2004, 81, 973–976. [CrossRef]
34. Hennet, M.L.; Yu, H.Y.; Combelles, C.M. Follicular fluid hydrogen peroxide and lipid hydroperoxide in bovine antral follicles of various size, atresia, and dominance status. J. Assist. Reprod. Genet. 2013, 30, 333–340. [CrossRef]
35. Ushio-Fukai, M.; Alexander, R.W. Reactive oxygen species as mediators of angiogenesis signaling: Role of NAD(P)H oxidase. Mol. Cell Biochem. 2004, 264, 85–97. [CrossRef]
36. Choi, J.; Jo, M.; Lee, E.; Choi, D. Induction of apoptotic cell death via accumulation of autophagosomes in rat granulosa cells. Fertil. Steril. 2011, 95, 1482–1486. [CrossRef]
37. Joshua McWhirt, M.S.; Sampath, D.; Newton, S.S. Effects of restraint stress on the regulation of hippocampal glutamate receptor and inflammation genes in female C57BL_6 and BALB_c mice. Neurobiol. Stress. 2019, 10, 100169. [CrossRef]
38. Geetha, A.; Shetty, B.H.; Upadhya, D.; Bates, A.; Attaluri, S.; Shuai, B.; Kodali, M.; Shetty, A.K. Chronic Oxidative Stress, Mitochondrial Dysfunction, Nrf2 Activation and Inflammation in the Hippocampus Accompany Heightened Systemic Inflammation and Oxidative Stress in an Animal Model of Gulf War Illness. Front. Mol. Neurosci. 2017, 10, 182. [CrossRef]
39. Zhao, L.; Li, H.; Wang, Y.; Zheng, A.; Cao, L.; Liu, J. Autophagy deficiency leads to impaired antioxidant defense via p62FOXO1/3 Axis. Oxidative Med. Cell. Longev. 2019, 2019, 2526314. [CrossRef] [PubMed]
40. Jeong, S.J.; Zhang, X.; Rodriguez-Velez, A.; Evans, T.D.; Razani, B. p62/SQSTM1 and Selective Autophagy in Cardiometabolic Diseases. Antioxid. Redox Signal 2019, 31, 458–471. [CrossRef]
41. Hennig, P.; Fenini, G.; Di Filippo, M.; Karakaya, T.; Beer, H.D. The Pathways Underlying the Multiple Roles of p62 in Inflammation and Cancer. Biomedicines 2021, 9, 707. [CrossRef]
[/registrados]