


| A lo largo de este artículo se efectúa una revisión sobre las últimas vacunas elaboradas para la prevención de la neumonía enzoótica porcina y se comenta su eficacia, después de una breve descripción de la enfermedad y su agente etiológico, y de un repaso a las vacunas tradicionales. |
LA ENFERMEDAD Y SU AGENTE ETIOLÓGICO
El Complejo Respiratorio Porcino (CRP) constituye una entidad multifactorial frecuente en la porcinocultura mundial que provoca pérdidas económicas cuantiosas derivadas de la conversión ineficaz del alimento, de la utilización de antibióticos en los tratamientos y de vacunas en la prevención.
No se trata de una zoonosis por lo que su transcendencia se ciñe a lo meramente pecuniario, con las únicas excepciones de algunas cepas de Pasteurella multocida, que pueden transmitirse raramente a través de las mordeduras o de secreciones nasales porcinas, o de Streptococcus suis, que podría ocasionar esporádicamente meningitis o septicemias humanas (Díez de los Ríos y cols., 2021).
CRP – LOS SOSPECHOSOS HABITUALES
| PATÓGENOS PRIMARIOS
Entre los patógenos primarios asociados al CRP se encuentran aquellos que ocasionan inmunosupresión, como el Virus del Síndrome Respiratorio y Reproductor Porcino (PRRSV), el Virus de la Influenza Porcina, el Virus de la Enfermedad de Aujeszky, Coronavirus, Circovirus y Mycoplasma hyopneumoniae, o que atacan a los fagocitos porcinos, como Actinobacillus pleuropneumoniae. |
PATÓGENOS SECUNDARIOS
Entre los patógenos secundarios u oportunistas figuran Bordetella bronchiseptica, Pasteurella multocida, Glaesserella parasuis o Streptococcus suis, que se aprovechan de la acción de los patógenos primarios (Assavacheep y Thanawongnuwech, 2022). |


| Si la enfermedad ocurre únicamente por efecto de los agentes primarios, los animales mueren o, alternativamente, el proceso remite en un tiempo razonable. Sin embargo, cuando se complica con los agentes oportunistas, el cuadro respiratorio se alarga, lo que origina unas pérdidas económicas de mayor envergadura (Gutiérrez Martín, 2014). |
MYCOPLASMA HYOPNEUMONIAE – ALLANANDO EL TERRENO PARA LOS OPORTUNISTAS

Como cualquier micoplasma, carece de pared bacteriana, pero puede fijarse al epitelio ciliado de la tráquea, bronquios y bronquiolos, ocasionando daños de consideración y facilitando el asentamiento de otras bacterias, principalmente A. pleuropneumoniae y P. multocida.
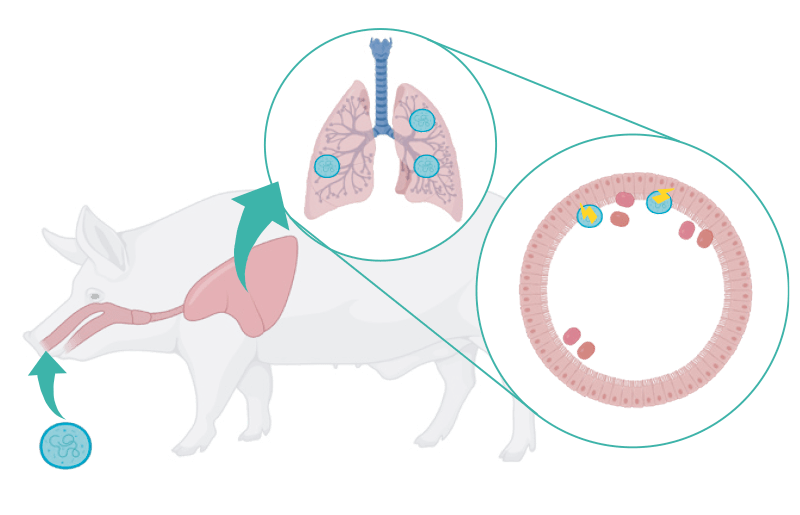

| 1 Fijación de M. hyopneumoniae al epitelio ciliado traqueal, bronquial y bronquiolar con generación de lesiones. | 2. Asentamiento de bacterias oportunistas. |
De distribución mundial, como factores predisponentes en las explotaciones intensivas cabe destacar:
- La ventilación insuficiente.
- Las fluctuaciones térmicas.
- La sobrepoblación porcina.
PATOGENIA Y SIGNOS CLÍNICOS
![]()

En su patogénesis resulta fundamental la acción sobre los macrófagos alveolares, que repercute en la reducción de su capacidad fagocítica, a lo que se suman la estimulación linfocitaria y de las citocinas proinflamatorias (Haesebrouck y cols., 2004; Gutiérrez Martín, 2014).
Resultan susceptibles los cerdos de todas las edades, y, como en cualquier proceso crónico, su mortalidad es escasa. Sin embargo, [registrados]tiene una morbilidad elevada asociada a:
- Tos improductiva.
- Asma.
- Anorexia y/o reducción de la ganancia de peso de los animales de cebo.
| ASPECTO MACROSCÓPICO
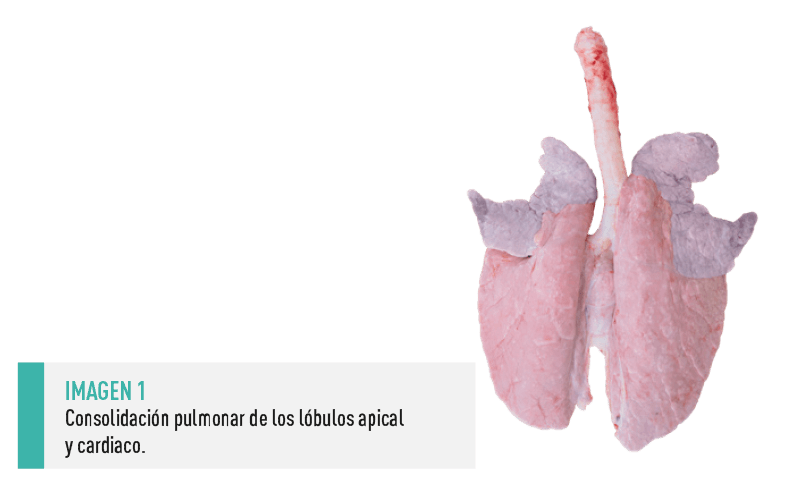
Su principal lesión macroscópica consiste en una consolidación pulmonar de los lóbulos apical y cardiaco, que suelen presentar una tonalidad púrpura-grisácea (Imagen 1). También puede resultar afectados los lóbulos intermedios y la parte anterior de los diafragmáticos (Maes y cols., 2008; Gutiérrez Martín, 2014; Quinn y cols., 2018). |
ASPECTO MICROSCÓPICO
Microscópicamente, se desarrolla una hiperplasia linforreticular peribroquial y perivascular como consecuencia de la respuesta inmunitaria (Haesebrouck y cols., 2004). |

DIAGNÓSTICO DE LA NEUMONÍA ENZOÓTICA
![]()
El diagnóstico clásico de la neumonía enzoótica se basa en el aislamiento e identificación del patógeno, lo que resulta difícil como todos los del género Mycoplasma. Por ello, se han diseñado otras alternativas más sensibles y eficaces, como la inmunofluorescencia a partir de las muestras pulmonares, o las técnicas de PCR.
LA DEMOSTRACIÓN DEFINITIVA EN LAS GRANJAS PORCINAS SUELE RESOLVERSE MEDIANTE UNA TÉCNICA ELISA
TRATAMIENTO Y PREVENCIÓN
![]()
Como antibióticos eficaces se han señalado la tilosina, la lincomicina o la tiamulina, aunque se han descrito resistencias frente a la primera, así como también frente a las fluoroquinolonas.



VACUNAS Y VACUNACIÓN FRENTE A LA NEUMONÍA ENZOÓTICA
La vacunación con formulaciones inactivadas comerciales representa una herramienta destacada en el control de la Neumonía Enzoótica, observándose una disminución parcial de las lesiones macroscópicas, así como una reducción de la carga de micoplasmas en el aparato respiratorio (Meyns y cols., 2006).
SE CALCULA QUE EN MUCHOS PAÍSES SE APLICAN VACUNAS EN MÁS DEL 70% DE LAS EXPLOTACIONES (Maes y cols., 2008)
No se conoce el mecanismo concreto de protección, pero se cita la inducción de inmunidad humoral en las mucosas y de inmunidad celular, alcanzándose la máxima eficacia varios meses después de su aplicación, no de modo inmediato (Haesebrouck y cols., 2004).
La ganancia diaria de peso asociada a la vacunación frente a este patógeno oscila entre el 2% y el 8%, con un índice de conversión del 2-5%, a lo que hay que añadir una disminución de la mortalidad y de los síntomas y lesiones.
No obstante, por lo general, esta protección no evita la colonización por M. hyopneumoniae, de cuyas cepas se han descrito variaciones genéticas y antigénicas destacadas, lo que complica el establecimiento de una protección vacunal óptima.
Para obtener resultados aceptables, la vacunación debe practicarse junto con otros factores de manejo (Haesebrouck y cols., 2004; Chae, 2011; Villarreal y cols., 2011).
Se han aplicado diferentes esquemas de vacunación, según la explotación y el patrón de infección.
![]()

![]()
![]()

En cualquier caso, el protocolo vacunal estará condicionado también por la presencia de otras enfermedades concurrentes o por el estrés (hacinamiento en el transporte, especialmente, a temperaturas elevadas, cambios en el manejo o la alimentación, etc.).
PESAR DE QUE ESTA CUESTIÓN HA SIDO MOTIVO DE DISCUSIÓN ENTRE LOS ESPECIALISTAS, SE HA SUGERIDO LA VACUNACIÓN TRES DÍAS ANTES DEL DESTETE COMO MOMENTO MÁS ADECUADO (Pieters y Sibila, 2017)


VACUNAS TRADICIONALES
Entre 2010 y 2017 se estudiaron diecinueve bacterinas que, en su mayoría, contenían la cepa J de M. hyopneumoniae, cepa tipo (Maes y cols., 2021) y, en menor número, se probaron también las cepas P, P-5722-3, BQ14 y DJ-166, concluyendo que las bacterinas no inducían niveles suficientes de anticuerpos protectores (Fish y cols., 2016).
A pesar de ello, la mayoría de ellas ha conseguido disminuir las lesiones neumónicas, aunque algunas ni eso, como la adyuvantada con un carbómero que, inoculada en cerdos por vía intramuscular, desarrolló el mismo grado de lesiones que el grupo control (Kristensen y cols., 2014).
Más recientemente, Matthijs y cols. (2019) ensayaron por vía intramuscular tres bacterinas basadas en la cepa F7.2C:
|
|
Todas estimularon una respuesta celular y redujeron los síntomas y lesiones, así como la presencia de ADN de M. hyopneumoniae en el pulmón. |
Kim y cols. (2021) evaluaron en cerdos la cepa 2940 inactivada potenciada con la endotoxina de la cepa J5 de E. coli inoculada en una única dosis por vía intramuscular.
El estudio de la expresión de las citoquinas en cerdos vacunados con una bacterina adyuvantada con una saponina puso de manifiesto la sobreexpresión de las relacionadas con una respuesta Th1 (IL-2, IL-12 y γ-interferón), lo que demostró la importancia de la inmunidad celular en la prevención de la neumonía enzoótica (Rodríguez y cols., 2023).
Entre 2015 y 2017, se valoraron siete vacunas formuladas con dos cepas atenuadas, la 168 y la RM48 (Tao y cols., 2019).
- Shen y cols. (2017) comprobaron que la vacuna atenuada que habían estudiado no favorecía una respuesta celular suficiente, al no estimular la síntesis de determinadas citoquinas. Años antes,
- Feng y cols. (2010) consiguieron una formulación atenuada con la cepa virulenta 168, aislada a partir de un cerdo enfermo, cuya virulencia había sido reducida mediante pases alternativos en el medio de Friis modificado y en el hospedador natural. Algo después, Feng y cols. (2013) incorporaron un mutante termosensible de la cepa LKR de M. hyopneumoniae en la formulación de una vacuna atenuada aplicada intranasalmente.
| Con carácter general, se admite que estas vacunas atenuadas garantizan únicamente una protección parcial, en el mejor de los casos. Además, siempre existe el problema de la virulencia residual y/o de la reversión de la cepa (Feng y cols., 2013). |

ANTÍGENO P97
![]()
La mayor parte de las investigaciones se refieren al antígeno P97, solo o en combinación con otras adhesinas, ya que es la principal proteína de membrana que se fija a los cilios del aparato respiratorio porcino. Se han realizado 16 estudios, entre ellos:

- Okamba y cols. (2007) indujeron una respuesta inmunitaria considerable cuando inocularon intramuscular e intranasalmente ratones con un adenovirus con la porción terminal carbohidratada de P97.
- Barate y cols. (2014) desarrollaron una vacuna recombinante de subunidades, adyuvantada con la subunidad B de la enterotoxina termolábil de Escherichia coli y con un aceite comercial, que probaron por vía intramuscular en ratón.

Su eficacia fue aceptable, puesto que la región R1 de la adhesina inducía una respuesta Th1, en tanto que su fragmento R2 generó una respuesta Th2.
- La vacuna formulada con parálogos recombinantes de las adhesinas ciliares P97/P102, potenciada con sendos adyuvantes comerciales (uno acuoso y otro oleoso) no rindió la eficacia pretendida por Woolley y cols. (2014), puesto que ni indujo inmunidad de mucosas ni pudo regularse la secreción de citoquinas proinflamatorias.
Como único resultado positivo, se desarrolló una respuesta inmunitaria sistémica aceptable.
- El estudio más reciente relacionado con P97 y otras proteínas inmunógenas (MHP418, MHP372, MHP199 y MHP0461) consistió en la construcción de tres proteínas quiméricas mediante métodos bioinformáticos que Indujeron una respuesta humoral de calidad en ratones, al tiempo que estimularon la secreción de TNF-α e IL-6.
Naturalmente, estos buenos resultados deben ser corroborados en cerdos (Rebouças Santos y cols., 2024).
ANTÍGENO P46
![]()
Se han realizado cinco experimentos en ratón relacionados con P46, antígeno de la superficie de la membrana, siempre combinado con al menos otras dos adhesinas.
- Destacamos el de de Oliveira y cols. (2017), quienes idearon una vacuna de ADN potenciada con una nanoemulsión comercial y observaron que P46 era reconocida por los sueros de los cerdos convalecientes.
- Liu y cols. (2023) construyeron una quimera multiantigénica denominada L9m6 (con P97R1, mhp390 y P46) y la probaron intramuscularmente en ratones.
Consiguieron una gran respuesta celular y una producción considerable de IgG e IgM. Todo ello sugirió el potencial vacunal de esta quimera que debería ser ratificado en el hospedador natural.
ANTÍGENO P36
![]()
La investigación de Zou y cols. (2011) es una de las tres que utiliza la adhesina P36 (localizada en el citosol) en un modelo ratón. Concretamente, este grupo recurrió al vector recombinante A. pleuropneumoniae SLW36 atenuado. Inoculada intramuscularmente, se generó una respuesta protectora frente a M. hyopneumoniae y A. pleuropneumoniae.
ANTÍGENOS P95 y P46
![]()
- La adhesina P95 fue incorporada en formulaciones vacunales de ADN y de subunidades por Galli y cols. (2012). Recurrieron al plásmido pcADN3 como vector de clonación y al hidróxido de aluminio como adyuvante, en un modelo murino.
Combinada con los antígenos P42 y P46 indujo resultados aceptables.
- Jorge y cols. (2014) prefirieron la proteína de choque térmico P42, que representa una parte de la proteína de 65 kDa expresada ante condiciones de estrés bacteriano. La adyuvantaron con un aceite comercial y fue inoculada por vía intramuscular a cerdos.
Desencadenó las dos ramas de la respuesta inmunitaria, humoral y celular, concluyéndose que P42 constituía un candidato antigénico prometedor para el desarrollo de vacunas.
- Chen y cols. (2003) también observaron una respuesta inmunitaria adecuada en ratones y cerdos cuando trabajaron con vacunas de ADN, concretamente, con el gen que expresaba esta misma proteína P42.
VÍAS DE ADMINISTRACIÓN INNOVADORAS
![]()
Hay vacunas cuya vía de administración no ha sido la habitual.
- La de Lin y cols. (2003) se basó en la administración oral de microesferas porosas de sílica con la cepa PRT-5 de M. hyopneumoniae, preparadas mediante un proceso de secado por pulverización conjunta.
Su efecto consistió fundamentalmente en la reducción de las lesiones neumónicas.
- Uno de los últimos experimentos recurrió al sondaje oral de lechones, a los que se les aplicaba una concentración proteica procedente de un lisado bacteriano sonicado de la cepa patógena 232 después de su autoclavado y ultracentrifugación.
Se logró una reducción de las lesiones pulmonares, así como la presencia de anticuerpos a la semana de la vacunación, que también protegían la mucosa respiratoria. Esta preparación ofreció resultados alentadores, aunque se precisan más estudios para concluir con seguridad su protección eficaz en producción porcina (Ferreira y cols., 2023).
- Mechler-Dreibi y cols. (2021) diseñaron una vacuna constituida por diversas proteínas encapsuladas por un material innovador, en forma de un complejo inmunógeno administrado por vía digestiva. Esta sustancia, denominada SBA-15, estaba formada por mesosporas de sílice de unos 10 nm de diámetro.
Preservaba los inmunógenos proteicos del pH ácido gástrico, pero se disolvía con el pH alcalino intestinal, lo que garantizaba la absorción de los inmunógenos vacunales en esta localización, sin haber sido dañados por las secreciones gástricas. Una vez más, esta formulación disminuyó las lesiones neumónicas y estimuló una inmunidad humoral aceptable de la mucosa digestiva, pero, como en otros estudios, estos resultados deben ser refrendados en un mayor número de animales.
- Canelli y cols. (2023) idearon un inmunógeno inactivado, potenciado con nanoesferas de quitosano, en forma de nanopartículas liofilizadas.
Se desarrolló una buena respuesta en la mucosa nasal a partir de la semana de la vacunación intranasal, así como la síntesis de gran cantidad de IgGs e IgAs y una secreción abundante de γ-interferón. Parecía convertirse de nuevo en otra opción de futuro alternativa a las vacunas parenterales convencionales.


VACUNAS MIXTAS
La inmunodepresión que suele surgir después de la neumonía enzoótica favorece la infección por agentes secundarios como el PCV2 o el PRRSV.
M. HYOPNEUMONIAE + PRRSV
![]()
- Una vacuna que combinó el PRRSV con M. hyopneumoniae es la de Drexler y cols. (2010), basada en el tipo europeo (atenuado) del PRRSV y en una bacterina de M. hyopneumoniae, que generó una protección aceptable frente a ambos microorganismos. A conclusiones similares llegaron Stricker y cols. (2013) y Bourry y cols. (2015).
M. HYOPNEUMONIAE + PCV2
![]()
- Herbich y cols. (2013) verificaron que la vacunación frente a M. hyopneumoniae y PCV2 resultaba tan eficaz como su inoculación por separado.
- Otro experimento llevado a cabo con estos dos mismos agentes ofreció buenos resultados en función de la ganancia diaria de peso, la viremia causada por el PCV2 y la gravedad de las lesiones (Kaalberg y cols. 2017).
M. HYOPNEUMONIAE + PCV2 + PRRSV
![]()
- La eficacia obtenida por Sipos y Sipos (2022) después de la administración de dos vacunas mixtas bivalentes que contenían PCV2 y PRRSV, una intramuscular y la otra intradérmicamente con un dispositivo sin aguja, fue más cuestionable, al conseguirse tan solo una estabilización de las lesiones pulmonares.
Sin embargo, del análisis de otras vacunas bivalentes comerciales integradas por estos dos mismos virus se obtuvieron resultados más alentadores (Yang y cols., 2021), como también del estudio de alguna trivalente que, junto con una cepa de M. hyopneumoniae, incluía los genotipos 2a y 2b del PCV (Um y cols., 2022; Yang y cols., 2022).
- Oh y cols. (2019) valoraron otra vacuna trivalente (PRRSV, PCV2 y M. hyopneumoniae). Aunque los resultados fueron buenos en términos de protección frente a estos tres agentes, su eficacia se resintió respecto a la aplicación por separado de las correspondientes vacunas monovalentes.
- La vacunación simultánea con la glucoproteína GP5 del PRRSV, la Cap del PCV2b y la adhesina P97 de M. hyopneumoniae también generaba una respuesta humoral aceptable en ratones (Roques y cols., 2013).
M. HYOPNEUMONIAE + B. BRONCHISEPTICA
![]()
- Como representación de una vacuna formulada por dos bacterias, podemos aludir a la que incorporó B. bronchiseptica y M. hyopneumoniae.
Yim y cols. (2017) comprobaron que la primera bacteria potenciaba en ratones la síntesis de IgGs específicas de la segunda.
| Como epílogo, se puede afirmar que la vacunación es utilizada en todo el mundo y las formulaciones más frecuentes han sido y siguen siendo las bacterinas intramusculares potenciadas con diferentes adyuvantes.
A pesar de los intentos de mejora explorados durante la última década, la eficacia vacunal continúa considerándose irregular debido a la gran variabilidad antigénica de las cepas circulantes, a los diferentes grados en la presión o a otros aspectos indeterminados de la respuesta inmunitaria, además de a diversos aspectos técnicos, como las condiciones inadecuadas de almacenamiento y/o administración o la falta de programas de vacunación. La inoculación intranasal de determinadas formulaciones ha producido resultados prometedores, que podrían convertirse en alternativas esperanzadoras, pero es aún largo el “camino experimental por recorrer”. |
BIBLIOGRAFÍA
Assavacheep, P., Thanawongnuwech, R. 2022. Porcine respiratory disease complex: Dynamics of polymicrobial infections and management strategies after the introduction of the African swine fever. Front Vet Sci. 9: 1048861.
Barate, A.K., Cho, Y., Truong, Q.I., Hahn, T.W. 2014. Immunogenicity of IMS 1113 plus soluble subunit and chimeric proteins containing Mycoplasma hyopneumoniae P97 C-terminal repeat regions. FEMS Microbiol. Lett. 352: 1213-220.
Bourry, O., Fablet, C., Simon, G., Marois-Crehan, C. 2015. Efficacy of combined vaccination against Mycoplasma hyopneumoniae and porcine reproductive and respiratory syndrome virus in dually infected pigs. Vet. Microbiol. 180: 230-236.
Canelli, E., Ferrari, L., Borghetti, P., Candela, F., Abiakam, N.S., Bianchera, A., Buttini, F., Magi, G.E., Sonvico, F., Martelli, P., Bettini, R. 2023. Nano-adjuvanted dry powder vaccine for the mucosal immunization against airways pathogens. Frontiers Vet. Sci. 10: 1116722.
Chae, Ch. 2011. Vaccination pigs against Mycoplasma hyopneumoniae infection: Failure to prevent transmission. Vet. J. 188: 7-8.
Chen, Y.L., Wang, S.N., Yang, W.J., Chen, Y.J., Lin, H.H., Shiuan, D. 2003. Expression and immunogenicity of Mycoplasma hyopneumoniae heat shock protein antigen P42 by DNA vaccination. Infect. Immun. 71: 1155-1160.
de Oliveira, N.R., Jorge, S., Gomes, G.K., Rizzi, C., Pacce, V.D., Collares, T.F., Monte, L.G., Dellagostin, O. 2017. A novel chimeric protein composed of recombinant Mycoplasma hyopneumoniae antigens as a vaccine candidate evaluated in mice. Vet. Microbiol. 201: 146-153.
Díez de los Ríos, J., Reynaga, E., García-González, M., Cámara, J., Ardanuy, C., Cuquet, J., Quesada, M.D., Navarro, M., Vilamala, A., Párraga-Niño, N., Quero, S., Romero, A., Benítez, R.M., Altimiras, J., Pedro-Botet, M.L. en representación del grupo de S. suis. 2021. Clinical and epidemiological characteristics of Streptococcus suis
infections in Catalonia, Spain. Front. Medicine 8: 792233.
Drexler, C.S., Witvliet, M.H., Raes, M., van de Laar, M., Eggen, A.A., Thacker, E.L. 2010. Efficacy of combined porcine reproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae vaccination in piglets. Vet. Record 166: 70-74.
Feng, Z., Shao, G., Liu, M., Wu, X., Zhou, Y., Gan, Y. 2010. Immune responses to the attenuated Mycoplasma hyopneumoniae 168 strain vaccine by intrapulmonic immunization in piglets. Agr. Sci. China 9: 423-431.
Feng, Z., Wei, Y., Li, G., Lu, X., Wan, X., Pharr, G., Wang, Z., Kong, M., Gan, Y., Bai, F., Liu, M., Xiong, Q., Wu, X., Shao, G. 2013. Development and validation of an attenuated Mycoplasma hyopneumoniae aerosol vaccine. Vet. Microbiol. 167: 417-424.
Ferreira, G.C., Sanches T.V.C., Mechler-Dreibi, M.L., Almeida, H.M.S., Storino, G.Y., Sonalio, K., Petri, F.A.M., Martins, T.S., Cides da Silva, L.C., Montassier, H.J., Sant’Anna, O.A., Fantini, M.C.A., de Oliveira, L.G. 2023. Efficacy evaluation of a novel oral silica-based vaccine in inducing mucosal immunity against Mycoplasma hyopneumoniae. Res. Vet. Sci. 158: 141-150.
Fisch, F.; Marchioro, S.B., Gomes, C.K., Galli, V., de Oliveira, N.R., Simionatto, S., Dellagostin, O.A., Mendonça, M., Moreira, A.N., Conceiçao, F.R. 2016. Commercial bacterins did not induce detectable levels of antibodies in mice against Mycoplasma hyopneumoniae antigens strongly recognized by swine immune system. Trials Vaccinol. 5: 32-37.
Galli, V., Simionatto, S., Marchioro, S.B., Fisch, A., Gomes, C.K., Conceicao, F.R., Dellagostin, O.A. 2012. Immunisation of mice with Mycoplasma hyopneumoniae antigens P37, P42, P46 and P95 delivered as recombinant subunit of DNA vaccines. Vaccine 31: 135-140.
García-Morante, B., Segales, J., Fraile, L., Llarden, G., Coll, T., Sibila, M. 2017. Potential use of local and systemic humoral immune response parameters to forecast Mycoplasma hyopneumoniae associated lung lesions. PLoS One 12: e0175034.
Gutiérrez Martín, C.B. 2014. La pleuroneumonía porcina en el contexto del Complejo Respiratorio Porcino. Conferencia leída en su recepción como académico correspondiente. Academia de Ciencias Veterinarias de Castilla y León. Servicio de Publicaciones. Universidad de León.
Haesebrouck, F., Pasmans, F., Chiers, K., Maes, D., Ducatelle, R., Decostere, A. 2004. Efficacy of vaccines against bacterial diseases in swine: what can we expect? Vet. Microbiol. 100: 255-268.
Herbich, E., Heissenberger, B., Ladinig, A., Griessler, A., Ritzmann, M., Weissenbacher-Lang, C., Hennig-Pauka, I., 2013. Field trial on the simultaneous vaccination against porcine circovirus type 2 (PCV2) and Mycoplasma hyopneumoniae. Tierartl. Prax. Ausg. G Grosstiere Nutztiere 41: 279-288.
Jorge, S., de Oliveira, N.R., Marchioro, S.B., Fisch, A., Gomes, C.K., Hartleben, C.P., Conceicao, F.R., Dellagostin, O.A. 2014. The Mycoplasma hyopneumoniae recombinant heat shock protein P42 induces an immune response in pigs under field conditions. Comp. Immunol. Microbiol. Infect. Dis. 37: 229-236.
Kaalberg, L., Geurts, V., Jolie, R. 2017. A field efficacy and safety trial in the Netherlands in pigs vaccinated at 3 weeks of age with a ready-to-use porcine circovirus type 2 and Mycoplasma hyopneumoniae combined vaccine. Porcine Health Manag. 3: 23.
Kim, S., Oh, T., Yang, S., Cho, H., Chae, Ch. 2021. Experimental evaluation of Mycoplasma hyopneumoniae bacterin against a Korean M. hyopneumoniae challenge. Can. J. Vet. Res. 85: 77-81.
Kristensen, Ch., Vinther, J., Svensmark, B., Bækbo, P. 2014. A field evaluation of two vaccines against Mycoplasma hyopneumoniae infection in pigs. Acta Vet. Scand. 56: 24.
Lin, J., Weng, C., Liao, C., Yeh, K., Pan, M. 2003. Protective effects of oral microencapsulated Mycoplasma hyopneumponiae vaccine prepared by co-spray drying method. J. Vet. Med. Sci. 65: 69-74.
Liu, W., Jiang, P., Song, T., Yang, K., Yuan, F., Gao, T., Liu, Z., Li, Ch., Guo, R., Xiao, S., Tian, Y., Zhou, D. 2023. A recombinant chimera vaccine composed of LTB and
Mycoplasma hyopneumoniae antigens P97R1, mhp390 and P46 elicits cellular immunologic response in mice. Vaccines 11: 1291.
Maes, D., Segales, J., Meyns, T., Sibila, M., Pieters, M., Haesebrouck, F. 2008. Control of Mycoplasma hyopneumoiae infections in pigs. Vet. Microbiol. 126: 297-309. Maes,
D., Boyen, F., Devriendt, B., Kuhnert, P., Summerfield, A., Haesebrouck, F. 2021. Perspectives for improvement of Mycoplasma hyopneumoniae vaccines in pigs. Vet. Res. 52: 67.
Matthijs, A.M.F., Auray, G., Boyen, F., Schoos, A., Michiels, A., García-Nicolás, O., Barut, G.T., Barnier-Quer, Ch., Jakob, V., Collin, N., Devriendt, B., Summerfield, A., Haesebrouck, F., Maes, D. 2019. Efficacy of three innovative bacterin vaccines against experimental infection with Mycoplasma hyopneumoniae. Vet. Res. 50: 91.
Mechler-Dreibi, M.L., Almeida, H.M.S., Sonalio, K., Martines, M.A.C., Petri, F.A.M., Zambotti, B.B., Ferreira, M.M., Storino, G.Y., Martins, T.S., Montassier, H.J., Sant’Anna, O.A., Fantini, M.C.A., de Oliviera, L.G. 2021. Oral vaccination of piglets against Mycoplama hyopneumoniae using silica SBA-15 as an adyuvant effectively reduced consolidation lung lesions at slaughter. Sci. Reports 11: 22377.
Meyns, T., Dewulf, J., de Kruif A., Calus, D., Haesebrouck, F., Maes, D. 2006. Comparison of transmission of Mycoplasma hyopneumoniae in vaccinated and
non-vaccinated populations. Vaccine 24: 7081-7086. Oh, T., Park, K.H., Yang, S., Jeong, J., Kang, I., Park, Ch., Chae, Ch. 2019. Evaluation of the efficacy of a trivalent vaccine mixture against a triple challenge with Mycoplasma hyopneumoniae, PCV2, and PRRSV and the efficacy comparison of the respective monovalent vaccines against a single challenge. BMC Vet. Res. 15: 342.
Okamba, F., Moreau, E., Bouh, C., Gagnon, C., Massie, B., Arella, M. 2007. Immune responses induced by replication-defective adenovirus expressing the C-terminal portion of Mycoplasma hyopneumoniae-P97 adhesin. Clin. Vaccine Immunol. 14: 767-774.
Pieters, M., Sibila, M. 2017. When is the best time to vaccinate piglets against Mycoplasma hyopneumoniae. Vet. Record July 1: 16-17.
Quinn, P.J., Markey, B.K., Leonard, F.C., FitzPatrick, E.S., Fanning, S., Hartigan, P.J. 2018. Microbiología y enfermedades infecciosas veterinarias. 2ª ed. Ed. Acribia, Zaragoza.
Rebouças Santos, M., Toledo, L.T., Bassi, E.J., Nascimento Porto, W.J., Costa Bressan, G., Moreira, M.A.S., Chang, Y.F., Silva Junior, A. 2024. Chimeric proteins of Mycoplasma hyopneumoniae as vaccine and preclinical model for immunological evaluation. Brazil J Microbiol. https://doi. org/10-1007/s42770-023-01240-7.
Rodríguez, F., Rosales, R.S., Ramírez, A.S., Poveda, J.B. 2023. Vaccination upregulates Th1 cytokines in the lung of pigs experimentally infected with Mycoplasma hyopneumoniae. Animals 13: 520.
Roques, E., Girard, A., Gagnon, C.A., Archambault, D. 2013. Antibody responses induced in mice immunized with recombinant adenovectors expressing chimeric proteins of various porcine pathogens. Vaccine 31:2698-2704.
Shen, Y., Hu, W., Wei, Y., Feng, Z., Yang, Q. 2017. The immune mechanism of Mycoplasma hyopneumoniae 168 vaccine strain through dendritic cells. BMC Vet. Res. 13: 285.
Shimoji, Y., Oishi, E., Muneta, Y., Nosaka, H., Mori, Y. 2003. Vaccine efficacy of the attenuated Erysipelothrix rhusiopathiae YS-19 expressing a recombinant protein of Mycoplasma hyopneumoniae P97 adhesin against mycoplasmal pneumonia of swine. Vaccine 2003, 21: 532-537.
Sipos, W., Sipos, S. 2022. Clinical efficacy of two novel, differentially administered (IM, ID) vaccines against Mycoplasma hyopneumoniae and PCV2 in swine under field conditions. Animals 12: 3467.
Stricker, T.M. Weissenbacher-Lang, C., Ritzmann, M., Ladinig, A., 2013. A study to assess the compatibility of simultaneous use of porcine reproductive and respiratory syndrome virus (EU-type) vaccine and Mycoplasma hyopneumoniae vaccine under field conditions. Berl. Munch. Tierztl. Wochenschr. 126: 189-196.
Tao, Y., Shu, J., Chen, J., Wu, Y., He, Y. 2019. A concise review of vaccines against Mycoplasma hyopneumoniae. Res. Vet. Sci. 123: 144-152.
Um, H., Yang, S., Oh, T., Cho, H., Park, K.H., Suh, J., Chae, Ch. 2022. A field efficacy trial of a trivalent vaccine containing porcine circovirus type 2a and 2b, and Mycoplasma hyopneumoniae in three herds. Vet. Med. Sci. 8: 578-590.
Villarreal, J., Meyns, T., Dewulf, J., Vranckx, K., Calus, D., Pasmans, F., Haesebrouck, F., Maes, D. 2011. The effect of vaccination on the transmission of Mycoplasma hyopneumoniae in pigs under field conditions. Vet. J. 188: 48-52.
Woolley, L.K., Fell, S.A., Gonsalves, J.R., Raymond, B.B.A., Collins, D., Kuit, T.A., Walker, M.J., Djordjevic, S.P., Eamens, G.J., Jenkins, Ch. 2014. Evaluation of recombinant Mycoplasma hyopneumoniae P97/P102 paralogs formulated with selected adjuvants as vaccines against mycoplasmal pneumonia in pigs. Vaccine 32: 4333-4341.
Yang, S., Ahn, Y., Oh, T., Cho, H., Park, K.H., Chae, Ch. 2021. Field evaluation of a single-dose bivalent vaccine of porcine circovirus type 2b and Mycoplasma hyopneumoniae. Vet. Med. Sci. 7: 755-765.
Yang, S., Lee, J.Y., Oh, T., Park K.H., Cho, H., Suh J., Min, K.D., Ham, H.J., Chae, Ch. 2022. Comparative growth performance of 3 types of combination vaccines containing porcine circovirus 2 and Mycoplasma hyopneumoniae under field conditions. Can. J. Vet. Res. 86: 93-101.
Yim, S.H., Hahn, T.W., Joo, H.G. 2017. Bordetella bronchiseptica antigen enhances the production of Mycoplasma hyopneumoniae antigen-specific immunoglobulin G in mice. J. Vet. Sci. 18: 327-332.
Zou, H.Y., Liu, X.J., Ma, F.Y., Chen, P., Zhou, R., He, Q.G. 2011. Attenuated Actinobacillus pleuropneumoniae as a bacterial vector for expression of Mycoplasma hyopneumoniae P36 gene. J. Gene Med. 13: 221-229.
Te puede interesar: Abordaje práctico al diagnóstico de la neumonía enzoótica porcina
[/registrados]