


El fósforo (P) es el segundo mineral más abundante en los animales después del calcio (Ca). En los cerdos, el 77% del P está en los huesos, mientras que el resto se distribuye en fluidos orgánicos y tejidos, participando en diversas funciones biológicas. Este artículo explora la importancia del fósforo en la salud y metabolismo de los cerdos, así como las implicaciones de su manejo dietético y ambiental.
EL PAPEL ESENCIAL DEL FÓSFORO EN LA SALUD Y METABOLISMO PORCINO
El fósforo (P) es el segundo mineral más abundante en los animales tras el calcio (Ca) en una relación fija P:Ca11, encontrándose aproximadamente el 80% en huesos y dientes, mientras que el 20% restante se distribuye en fluidos orgánicos y tejidos, participando en diversas funciones biológicas.
| En el cerdo, aproximadamente el 77% del P se encuentra en los huesos, en comparación con el 99% del Ca. La proporción de P en los huesos varía según el estado del hueso, constituyendo el 18% de las cenizas del hueso, el 10% del hueso seco y libre de grasa, y el 4,5% del hueso húmedo. |
En el líquido extracelular, aproximadamente el 30% del P, circula como ortofosfato, PO43– y colabora en el mantenimiento de la presión osmótica, el equilibrio ácido-base, la actividad neuronal y el apetito6.
El P juega un papel vital en una serie de funciones metabólicas:
- Utilización y transferencia de energía a través de AMP, ADP, ATP17 y fosfocreatina12 implicándose en la gluconeogénesis.
- Transporte de ácidos grasos.
- Síntesis de proteínas.
- Actividad de la bomba de sodio y potasio.
- Control del apetito y eficiencia de conversión de los alimentos28.
Por otro lado, el P forma parte de las moléculas de ARN y ADN, siendo esencial para el crecimiento y diferenciación celular, y, asociado a lípidos, forma parte de los fosfolípidos que contribuyen a la fluidez de las membranas celulares y a la mielinización de los nervios28.
En general, los depósitos de P en el organismo se pueden imaginar como un pequeño depósito circulante (fluidos corporales) para funciones vitales a corto plazo y un gran depósito en el esqueleto.
| El esqueleto, a través de la deposición y resorción continuas, proporciona capacidad de amortiguamiento finita para mantener niveles constantes en los tejidos blandos. |
Las necesidades de P se estiman junto con las del Ca y no pueden estimarse de forma separada12. No obstante, en este artículo se obviarán las implicaciones del Ca por razones didácticas.
El P elemental es altamente reactivo y se transforma en fosfato al exponerse al aire, siendo esta la forma nutricionalmente relevante de P para plantas y animales, obteniéndose principalmente de la roca fosfórica, un recurso no renovable, finito.
UN MANEJO INADECUADO DEL P EN LA ALIMENTACIÓN ANIMAL PUEDE RESULTAR EN UNA BAJA PRODUCTIVIDAD DEBIDO A UNA DEFICIENCIA O UN EXCESO DE P
Un suministro excesivo puede causar lixiviación del P a través de vías fluviales, provocando graves consecuencias ecológicas, como la eutrofización de las aguas que favorece el crecimiento de algas tóxicas fijadoras de nitrógeno o cianobacterias30.
Debido a la regulación hormonal, los valores de P en plasma no siempre reflejan el estatus mineral del animal, ya que fluctúan ampliamente en animales sanos, y simplemente reflejan el equilibrio entre las entradas y salidas de P28,29. Pese a ello, se consideran valores normales28:
- 2,6-3,2 mmol L-1 para animales con pesos inferiores a 25 kg.
- 2,3-2,6 mmol L-1 para animales con pesos superiores a 25 kg.
El P aumenta después de la ingestión y se reduce por el estrés.
| A este respecto, un estudio realizado por Bautista et al. (2010) aplicando P por vía intramuscular a cerdos a los 3 días de vida y al destete confirmó la relación entre estrés, fosfatemia y tasa de crecimiento, ya que los animales obtuvieron mejores ganancias de peso (+58 g/d) y llegaron al peso de sacrificio con 8,5 kg más de peso vivo. |
INOSITOL, ÁCIDO FÍTICO, FITATOS Y FITINA
La relación entre el inositol, ácido fítico y el fitato en el P de las plantas es fundamental en la nutrición y bioquímica vegetal.
INOSITOL
El inositol (ciclohexano-1, 2, 3, 4, 5, 6-hexol; C6H12O6) es un alcohol poliol del ciclohexano y componente principal del ácido fítico.
EL ISÓMERO MÁS COMÚN DEL INOSITOL ES EL MIOINOSITOL, UNA FORMA BIOLÓGICAMENTE ACTIVA SIMILAR A LA GLUCOSA
La fosforilación del mioinositol da lugar a ácido fítico, un proceso eficiente para almacenar grandes cantidades de P en una molécula compacta.
El mioinositol es abundante en granos de cereal y subproductos de soja.
Se absorbe eficientemente en el intestino delgado de los cerdos y se distribuye a varios tejidos, incluidas las células del hígado y músculos, desempeñando un papel crucial en funciones celulares y biológicas como:
|
ÁCIDO FÍTICO
El ácido fítico o hexafosfato de inositol (PI6) es una molécula compuesta por un anillo de inositol al que están unidos 6 grupos fosfato, siendo la principal forma de almacenamiento de P en las semillas y los granos, representando hasta el 90% del P total en las semillas26.
| El ácido fítico se une débilmente a cationes bi- y trivalentes en condiciones ácidas del estómago y precipita como fitato en el pH neutro del intestino delgado, inhibiendo la absorción intestinal de oligoelementos, considerándose por ello un antinutriente. |
FITATOS
Los fitatos son [registrados]las sales de ácido fítico que se forman al unirse a cationes, proteínas, aminoácidos y almidón para aumentar su estabilidad química, siendo esenciales para la germinación de las semillas al formar parte de hormonas como el ácido abscísico y las giberelinas necesarias para este proceso26.
Son solubles a pH ácido y se recombinan a pH básico, una característica fundamental en la nutrición animal.
El P ligado al fitato (282 g kg-1 de P en el fitato22), denominado comúnmente P fítico, es solo parcialmente degradable en los cerdos debido a:
|
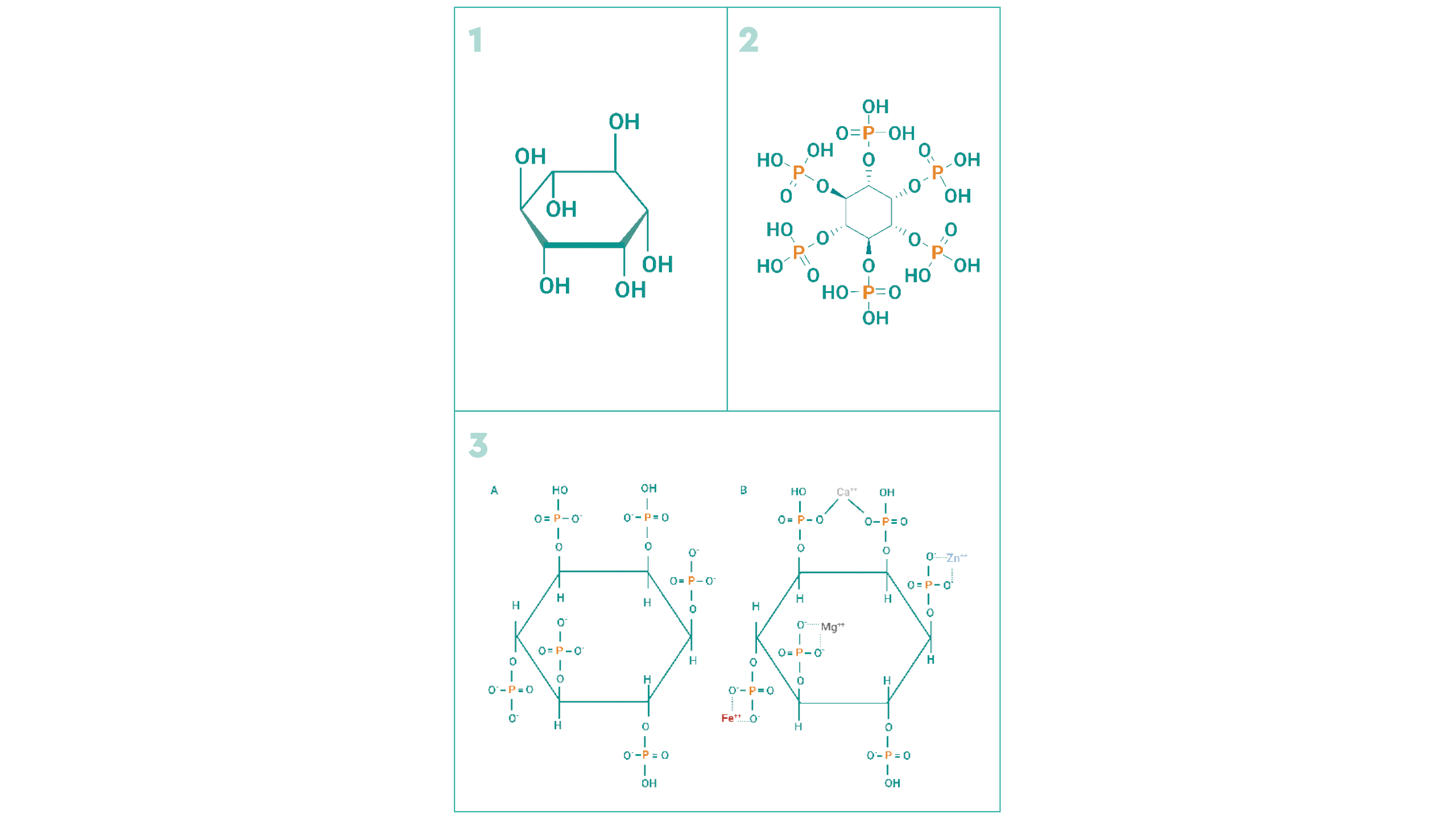

| FIGURA 1 Estructura del inositol (1), ácido fítico (2) y fitato11 (3A) a pH neutro y (3B) quelado con diferentes cationes. |
LA CONCENTRACIÓN DE FITATO EN LA DIETA DETERMINA LA MAGNITUD DE LA RESPUESTA DEL ANIMAL A LA SUPLEMENTACIÓN DE FITASA EXÓGENA
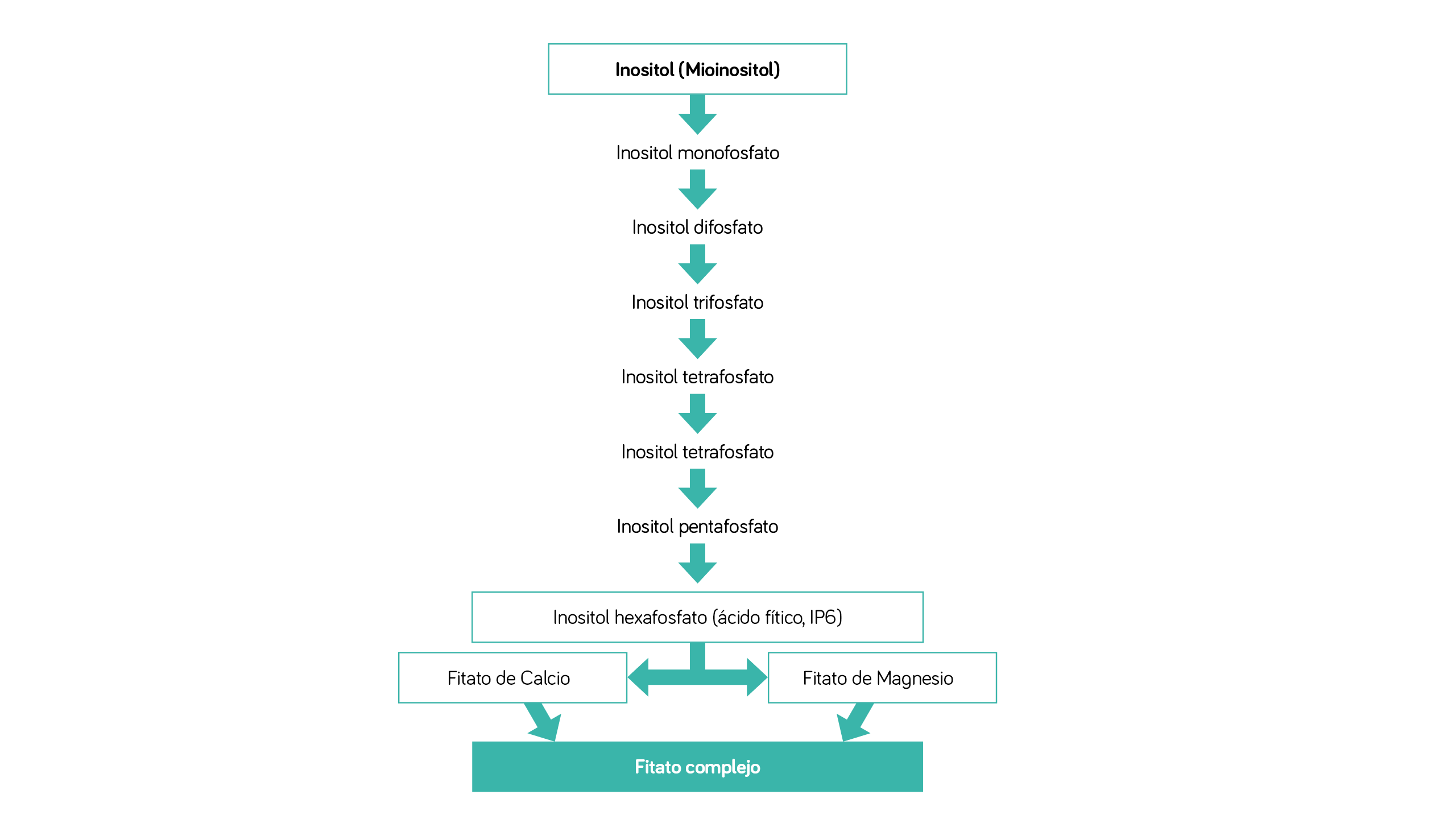

| FIGURA 2 Metabolismo del inositol, ácido fítico y fitatos en las plantas. |
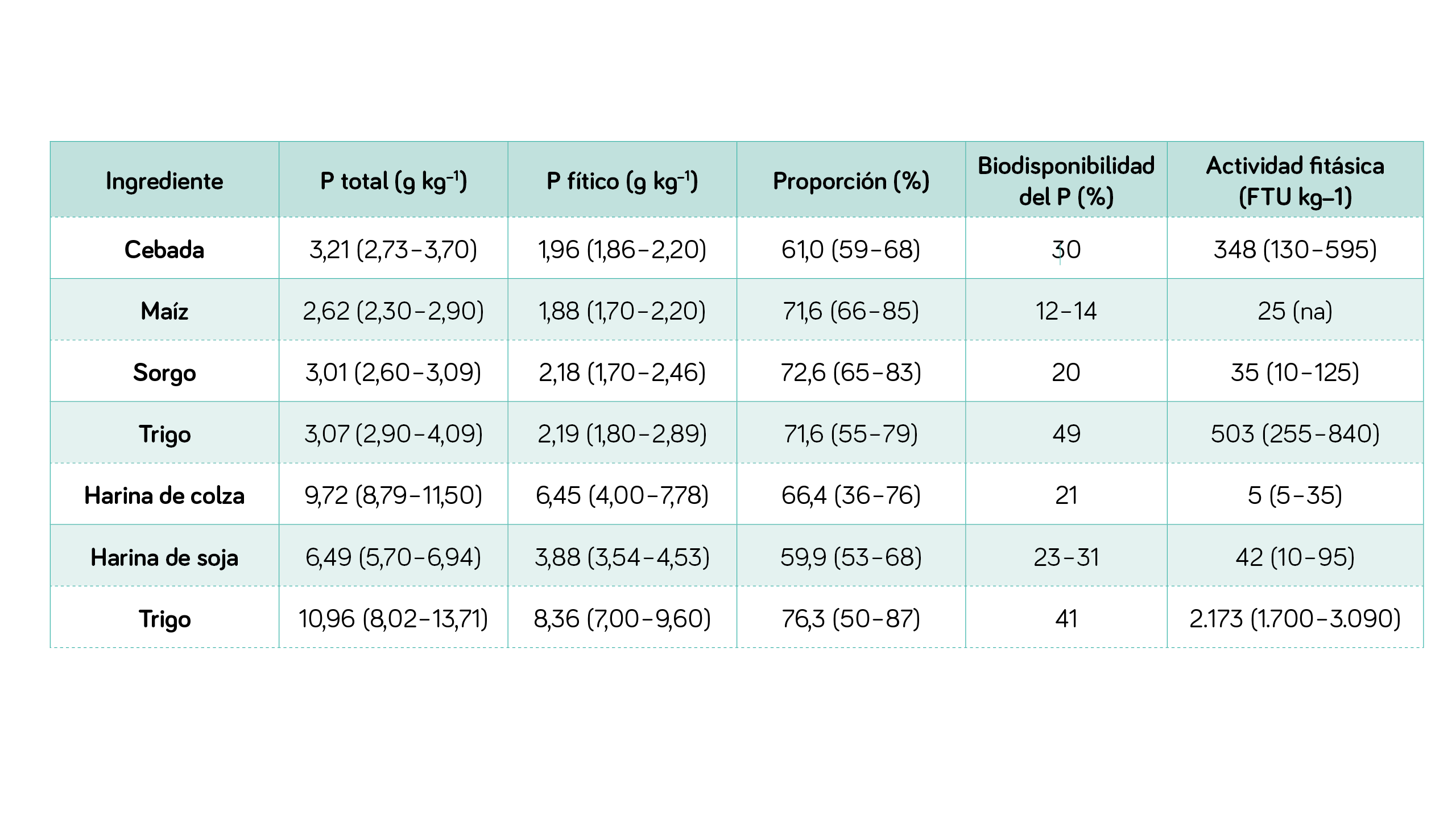

| TABLA 1 Valores de P total, fítico, disponibilidad del P y actividad fitásica intrínseca de algunas materias primas (Modificado de Selle y Ravindran, 2008). |
| La fitina es una mezcla de ácido fítico y sus sales de Ca, magnesio y potasio, siendo una forma especial de fitato presente en las plantas. No todos los fitatos son fitina, ya que se trata de una forma específica de fitato. |
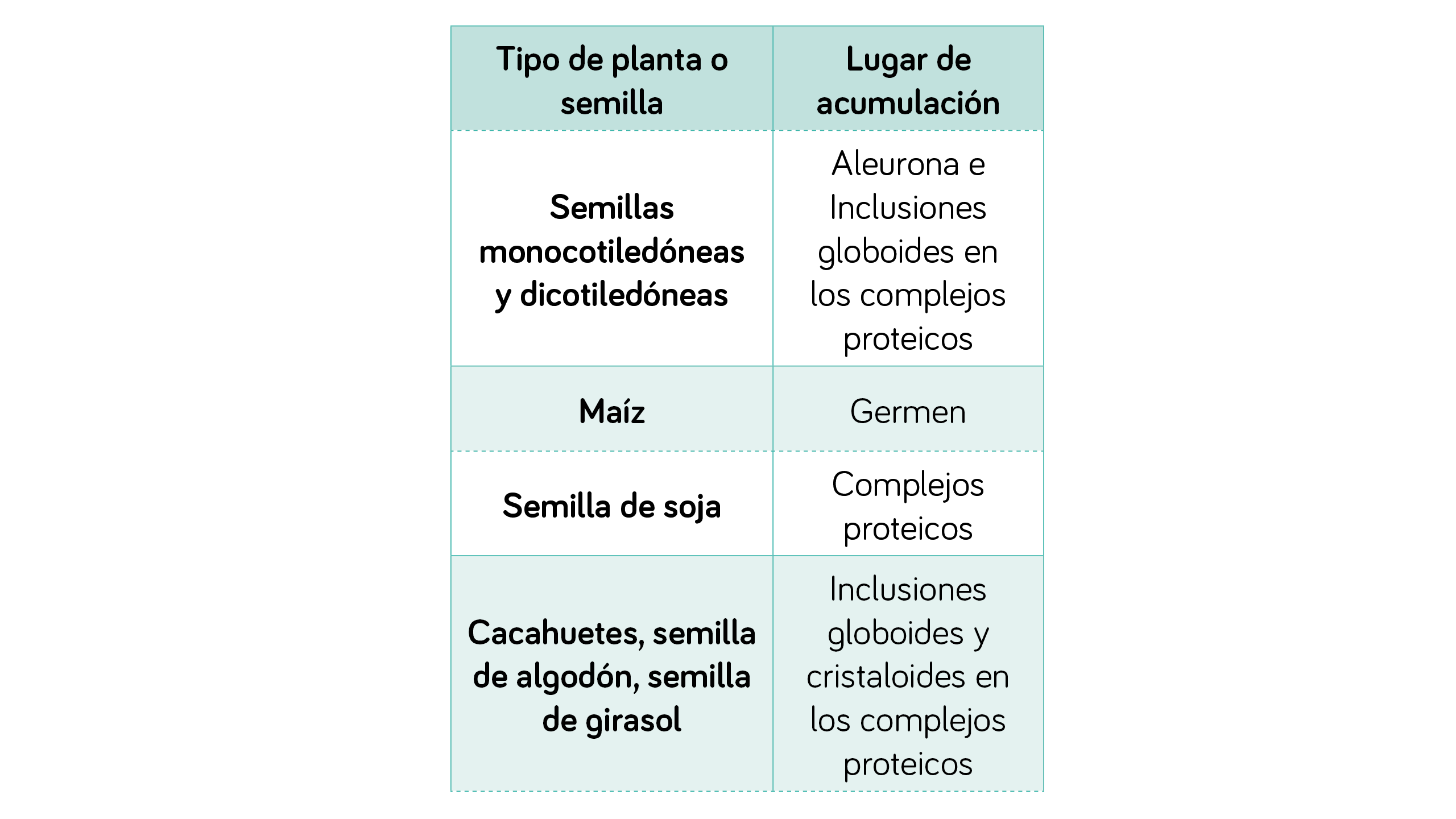

| TABLA 2
Acumulación de fitina en las plantas7. |
Se han desarrollado variedades de cereales y oleaginosas para reducir la concentración de fitato y aumentar la biodisponibilidad del P en los piensos.
| Por ejemplo, la digestibilidad del P en el maíz bajo en fitato fue aproximadamente 26 puntos porcentuales mayor que en el maíz normal, y la biodisponibilidad del P en la harina de soja baja en fitato fue de 12 a 26 puntos porcentuales mayor que en la harina de soja convencional31. |
FÓSFORO FÍTICO Y FÓSFORO NO FÍTICO
| FÓSFORO FÍTICO
El fósforo fítico (PP, Phytic Phosphorus), proviene del ácido fítico y de los fitatos, siendo una forma de almacenamiento de P en plantas, especialmente en semillas y granos, constituyendo hasta el 60-80% del P total en las plantas.
|

| FÓSFORO NO FÍTICO
El fósforo no fítico (NPP, Non Phytic Phosphorus) está presente en formas más fácilmente biodisponibles, como fosfatos inorgánicos y ésteres fosfóricos.
Sin embargo, esto no implica que, en todas las materias primas, el NPP sea totalmente digestible.
No todo el NPP es 100% disponible, por ello, estos conceptos serán analizados al discutir la digestión, absorción y uso del P en cerdos y las nuevas formas de analizar y medir la digestibilidad del P. |

METABOLISMO DEL FÓSFORO EN CERDOS31
DIETAS SIN FITASA
![]()

La hidrólisis del ácido fítico y fitatos ocurre de forma gradual, produciendo fosfatos libres y varios productos intermedios que pueden ser absorbidos en pequeña medida.
![]()

DIETAS CON FITASA
![]()
El principal sitio de hidrólisis del fitato por fitasas suplementadas es el estómago, ya que el pH gástrico se ajusta mejor a los óptimos de pH de la mayoría de las fitasas comerciales en comparación con el pH intestinal.
| La mayor diferencia en la degradación del ácido fítico atribuible a la suplementación de fitasa se encuentra en el estómago y el duodeno, manteniéndose esta diferencia a lo largo del intestino delgado antes de la actividad microbiana adicional en el intestino grueso. |
ABSORCIÓN DE FÓSFORO
El duodeno y el yeyuno son los principales sitios de absorción del P en los cerdos y otras especies monogástricas, y ocurre gracias a transportadores específicos de fosfato/sodio en el intestino delgado, facilitando un proceso activo y transcelular10.
![]()
En lechones la absorción puede alcanzar el 97%6,28.
| La solubilidad de las fuentes de P influye en su absorción, siendo el P inorgánico más fácilmente absorbible en comparación con las fuentes de P de origen vegetal. |
LAS FITASAS EXÓGENAS AYUDAN A LIBERAR EL P DE LOS FITATOS
El intestino grueso también contribuye a la absorción del P en los cerdos14,15, ya que, aunque en menor medida que el intestino delgado, la expresión de transportadores de fosfato/sodio en el colon sugiere una absorción significativa de P en esta sección. Además, el intestino grueso puede desempeñar un papel importante en el reciclaje del P endógeno secretado en la parte superior del tracto gastrointestinal.
La absorción neta de P en el intestino grueso puede depender de la fuente de carbohidratos en la dieta:
- La celulosa y la pectina se asocian a una secreción neta de P.
- El almidón se asocia a una absorción neta de P.
En dietas prácticas, se ha observado una mayor absorción de P en el intestino grueso en comparación con dietas semi-purificadas5.
| En ratas, la absorción de P se produce por vía transcelular (78%) o paracelular (22%) de forma activa y pasiva cuando las concentraciones luminales de P son muy altas, y se presume que este modelo es similar en cerdos8. |
EXCRECIÓN DE FÓSFORO
Una vez absorbido, el P viaja por la sangre asociado a fosfolípidos o como P inorgánico alcanzando los diferentes tejidos y, según las necesidades, se retendrá en los tejidos o se excretará a través de la leche, las heces o la orina, siendo esta última la principal vía de eliminación. La regulación del metabolismo del P involucra la interacción cooperativa del intestino, los huesos y el riñón, de la paratohormona, el calcitriol y la calcitonina18,28.
![]()
| La eliminación de estiércol con alto contenido de P plantea preocupaciones ambientales debido al exceso de P en el estiércol comparado con el requerido por los cultivos, lo que puede llevar a su acumulación en el suelo y su eventual movimiento durante la erosión y el escurrimiento superficial del agua. |
HOMEOSTASIS DEL FÓSFORO
La homeostasis del P se regula mediante la absorción intestinal, la movilización de P desde los huesos y la excreción renal a través de varios mecanismos hormonales y metabólicos:
![]()
![]()
| Se activa a través de su conversión en el hígado a 25-hidroxicolecalciferol* y luego en los riñones a 1,25-dihidroxicolecalciferol (calcitriol)**, su forma más activa. |
![]()
![]()
| *25-hidroxicolecalciferol: metabolito circulante principal de la vitamina D3 que se forma en el hígado, actuando como un precursor que se convierte en los riñones en la forma activa de vitamina D3. **1,25-dihidroxicolecalciferol (calcitriol): metabolito más activo de la vitamina D3 que se produce en los riñones, aumentando la absorción de P en el intestino delgado, facilitando la liberación de P desde los huesos y reduciendo la excreción renal de P. El 24,25-dihidroxicolecalciferol, un metabolito menos activo de la vitamina D3, también se produce en los riñones. Su función precisa no está completamente aclarada, pero se considera menos eficiente en la regulación de la homeostasis del P comparado con el calcitriol. |
DISPONIBILIDAD DEL FÓSFORO
P DISPONIBLE, BIODISPONIBILIDAD O DISPONIBILIDAD DEL P
La biodisponibilidad (P disponible) es un concepto abstracto para describir el grado en que el P en un alimento es absorbido, metabolizado y utilizado por el animal, pero se ha utilizado sin la debida prudencia en muchas publicaciones27.
![]()
Para estimar las necesidades de P en los cerdos, se ha utilizado ampliamente el concepto de P disponible y, como se ha comentario anteriormente:
- El P fítico (PP) se consideraba no disponible.
- El P no fítico (NPP) se consideraba disponible para el cerdo.
Tradicionalmente, el P disponible se determinaba mediante el Valor Biológico Relativo (RBV, Relative Biological Value), evaluando la resistencia ósea, el peso de ceniza ósea y el porcentaje de ceniza ósea en cerdos con diversas fuentes de P y comparándolos con una fuente estándar.
| La RBV de una fuente estándar de P asigna un valor del 100% y la biodisponibilidad del P en otras materias primas se estima como un porcentaje relativo utilizando el método de la razón de pendientes (Slope Ratio Method), como se describe en Petersen et al. (2011). Este método es útil para comparar la biodisponibilidad de diferentes fuentes de P, pero presenta desventajas. |
LA RBV DEL P NO ES ADITIVA EN DIETAS MIXTAS PARA CERDOS23
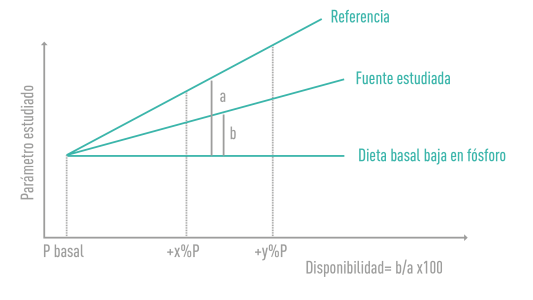

| FIGURA 3
Descripción del método de la razón de pendientes o Slope Ratio Method. |
La disponibilidad del P depende de varios factores3,16,19,18:
- La variable respuesta escogida para su evaluación, como el peso corporal del cerdo, las cenizas del hueso o la dureza a la rotura del fémur.
- El fosfato inorgánico escogido como referencia.
- La composición de la dieta.
- La relación entre el Ca y el P, que varía entre las diferentes materias primas.
- Edad del cerdo o su desarrollo (lechón, finalizador o hembra).
- Duración del ensayo.
Parafraseando a Leske y Coon (2002) cuando se refieren al P disponible al calcularlo en aves: “El valor biológico (biodisponibilidad de P) solo describe el potencial de utilización del P de una materia prima cuando se compara con un estándar de referencia para el P contenido en los fosfatos usados en el pienso. La cantidad real de P retenido por el ave por unidad de P en la materia prima no se determina”.
HOY EN DÍA, SE ESTIMAN LAS NECESIDADES DEL P EN TÉRMINOS DE DIGESTIBILIDAD Y SE HA DEJADO DE USAR LA DISPONIBILIDAD
DIGESTIBILIDAD DEL FÓSFORO
La estimación de la digestibilidad puede realizarse mediante dos métodos complementarios: la estimación factorial y la estimación empírica que valida la estimación factorial.
La estimación factorial se basa en estimar las necesidades para:
- El mantenimiento (reposición de las pérdidas endógenas inevitables o basales en las heces y la orina).
- La retención de P en el cuerpo y en los productos de la concepción.
- La excreción de P en la leche.
+
| La eficiencia con la que el P digestible se utiliza para estos procesos. |
Para estimar las necesidades de mantenimiento, primero se deben calcular las pérdidas endógenas de P, que pueden dividirse en basales y específicas.
| PÉRDIDAS ENDÓGENAS BASALES
Pérdidas asociadas a las características inherentes a las necesidades del animal, incluidas en las necesidades nutricionales.
|
| PÉRDIDAS ENDÓGENAS ESPECÍFICAS
Pérdidas asociadas a la fibra dietética, considerándose una característica de los ingredientes alimentarios que se contabilizan implícitamente en el coeficiente de digestibilidad.
|
DIGESTIBILIDAD DEL P
![]()
En teoría, la digestibilidad del P en un ingrediente puede calcularse multiplicando su biodisponibilidad relativa (RBV) por la digestibilidad del P en el P mineral estándar con el que se compara, aunque esta conversión ha sido cuestionada por estudios comparativos.
Para medir la digestibilidad del P, se requieren varias decisiones operativas:
- El uso de un marcador o la comparación cuantitativa clásica entre la entrada y salida.
- La elección entre el muestreo ileal y fecal.
ELECCIÓN ENTRE MUESTREO ILEAL Y FECAL
![]()
Se ha demostrado que el intestino grueso no juega un papel importante en la digestión del P y no hay diferencia en la digestibilidad verdadera del P entre los niveles ileal y total del tracto digestivo.
![]()
| La secreción o absorción de P en el intestino grueso no se puede integrar fácilmente en el sistema de digestibilidad actual, y su influencia en la formulación precisa de dietas en condiciones prácticas puede ser insignificante. |
DIGESTIBILIDAD APARENTE DE P O ATTD DEL P
![]()
El término “aparente” de la digestibilidad del P (ATTD, Apparent Total Tract Digestibility) hace referencia al cálculo en el que, tanto el P no digerido del alimento como la pérdida endógena del cerdo, han sido deducidos del suministro dietético de P.
Esta medida toma en cuenta el fósforo ingerido y el fósforo excretado en las heces, sin diferenciar entre el fósforo que proviene del alimento y el fósforo que se pierde endógenamente (es decir, el fósforo que el animal excreta naturalmente, independientemente del fósforo en el alimento).
ATTD del P% = [(P ingerido – P excretado)/P ingerido] x 100
Algunos estudios han encontrado que los valores de ATTD del P medidos para los ingredientes no siempre son aditivos cuando se utilizan en la formulación de dietas para cerdos. Por ejemplo:
- Los valores de ATTD para el P en el maíz varían significativamente en estudios con cerdos de diferentes pesos1,9,25.
- En cambio, de ATTD del P en el fosfato monocálcico no cambia con el aumento de su tasa de inclusión.
- Los valores la ATTD en la harina de soja y harina de colza tampoco varían con el nivel de inclusión19.
| La pérdida endógena de P de los cerdos representa diferentes proporciones del suministro de P debido a las tasas variables de inclusión de las diferentes materias primas del pienso, lo que desafía la aditividad de la digestibilidad aparente del P, un problema que se minimiza al usar el sistema de digestibilidad estandarizada del P. |
DIGESTIBILIDAD ESTANDARIZADA DEL P O STTD DEL P
![]()
El término “estandarizado” (STTD, Standardized Total Tract Digestibility) hace referencia al cálculo en el que, tanto el P no digerido del alimento como la pérdida endógena específica de P del cerdo han sido deducidos del suministro de P del alimento, mientras que la inevitable pérdida endógena basal de P se considera un factor de las necesidades del cerdo.
Las pérdidas endógenas basales son las pérdidas inevitables de fósforo que ocurren en el tracto digestivo del cerdo, que no están relacionadas con el tipo de alimento. Se consideran una constante.
La STTD se obtiene ajustando la digestibilidad aparente (ATTD) para incluir únicamente las pérdidas endógenas basales, y no las pérdidas endógenas totales (que incluirían pérdidas adicionales específicas del tipo de alimento).
STTD del P (%) = [(P ingerido – (P excretado – Pérdidas endógenas basales de P))/P ingerido] × 100
- El NRC18 calculó la STTD del P para las diferentes materias primas utilizando pérdidas endógenas basales de 190 mg/kg de materia seca consumida para corregir los valores de ATTD del P. Sin embargo, Stein et al. (2011) en sus cálculos usaron el valor de 200 mg/kg para corregir los valores de ATTD del P.
- Almeida y Stein (2010) demostraron indirectamente que los valores de STTD para el P son aditivos, ya que no observaron diferencias significativas entre cuatro dietas formuladas utilizando valores de STTD de P.
- She et al. (2018) demostraron que la STTD de P en maíz, harina de soja y harina de colza es más aditiva que sus equivalentes de ATTD en cerdos.
SEGÚN EL NRC18, EL USO DE VALORES DE STTD DE P EN LA FORMULACIÓN PRÁCTICA DE DIETAS PERMITE LOGRAR ADITIVIDAD ENTRE LOS INGREDIENTES DEL ALIMENTO Y FORMULAR DIETAS DE MANERA MÁS PRECISA EN COMPARACIÓN CON EL USO DE VALORES DE ATTD DE P
DIGESTIBILIDAD VERDADERA DEL P O TTTD DEL P
![]()
El término “verdadero” de la digestibilidad del P (TTTD, True Total Tract Digestibility) hace referencia a la capacidad real de un alimento para proporcionar P, según el cálculo en el que solo el P no digerido derivado del alimento ha sido deducido del suministro de P en el alimento.
Se han utilizado diferentes métodos para medir la digestibilidad verdadera del P:
- Fan et al. (2001) determinaron la pérdida endógena específica de P y los valores de digestibilidad verdadera del P en ingredientes vegetales utilizando la técnica de regresión y el método del índice.
- Petersen et al. (2011) estimaron las pérdidas endógenas específicas de P y la digestibilidad verdadera de P en fuentes inorgánicas de P basándose en una dieta libre de P y, junto a otros autores, probaron la aditividad de los valores de TTTD del P en diferentes materias primas vegetales.
| El principal desafío de la TTTD es la dificultad para medir la pérdida endógena específica de P asociada a cada ingrediente y considerarla como parte de los requerimientos del cerdo. En cambio, el sistema de STTD, que corrige el P digestible aparente por la pérdida endógena basal, parece ser la opción más práctica. |
ESTAS METODOLOGÍAS ESTIMAN NECESIDADES, PERO NO LAS RESPUESTAS DEL CERDO, COMO DEFICIENCIAS TRANSITORIAS O RETENCIONES COMPENSATORIAS QUE OCURREN EN LA PRÁCTICA
La cantidad precisa de P en la dieta de un cerdo depende de la interacción entre los objetivos productivos (rentabilidad, bienestar animal y protección medioambiental) y los criterios de respuesta (crecimiento diario, excreción de P al medio ambiente o mineralización ósea).
Por ello, las técnica multiobjetivo de maximización o minimización requieren modelos matemáticos mecanísticos complejos, como el presentado en Lautrou et al. (2020 y 2021).
| En conclusión, el fósforo es esencial para el metabolismo y la salud de los cerdos, desempeñando funciones cruciales en el crecimiento óseo, la síntesis de proteínas y la actividad enzimática. |
Te puede interesar: Estrategias de bioseguridad frente a virus porcinos en los piensos
| BIBLIOGRAFÍA 1. Ajakaiye, A,, M, Z, Fan, T, Archbold, R, R, Hacker, C, W, Forsberg, y J, P, Phillips, 2003, «Determination of true digestive utilization of phosphorus and the endogenous phosphorus outputs associated with soybean meal for growing pigs1», Journal of Animal Science 81 (11): 2766- 75, https://doi,org/10,2527/2003,81112766x.2. Almeida, F, N,, y H, H, Stein, 2010, «Performance and phosphorus balance of pigs fed diets formulated on the basis of values for standardized total tract digestibility of phosphorus», Journal of Animal Science 88 (9): 2968-77, https://doi.org/10,2527/jas,2009-2285.3. Baker, David H,, y Hans H, Stein, 2013, «Bioavailability of Minerals and Vitamins in Feedstuffs», En Sustainable Swine Nutrition, 341-64, John Wiley & Sons, Ltd, https://doi.org/10.1002/9781118491454.ch15.4. Barlet, J,, M, Davicco, y V, Coxam, 1995, «Physiology of intestinal absorption of phosphorus in animals,» Reproduction, nutrition, development 35 5:475-89,5. Baumgärtel, Tina, Barbara U, Metzler, Rainer Mosenthin, Ralf Greiner, y Markus Rodehutscord, 2008, «Precaecal and postileal metabolism of P, Ca and N in pigs as affected by different carbohydrate sources fed at low level of P intake», Archives of Animal Nutrition 62 (3): 169-81, https://doi.org/10.1080/17450390802028047. 6. National Research Council, 2005, Mineral Tolerance of Animals: Second Revised Edition, 2005. https://doi.org/10.17226/11309. 7. Eeckhout, W,, y M, De Paepe, 1994, «Total phosphorus, phytate-phosphorus and phytase activity in plant feedstuffs», Animal Feed Science and Technology 47 (1): 19-29. https://doi.org/10.1016/0377-8401(94)90156-2. 8. Eto, Nobuaki, Mikio Tomita, y Masahiro Hayashi, 2006, «NaPi-mediated Transcellular Permeation is the Dominant Route in Intestinal Inorganic Phosphate Absorption in Rats», Drug Metabolism and Pharmacokinetics 21 (3): 217-21, https://doi.org/10.2133/dmpk.21,217. 9. Fan, Ming Z,, Tania Archbold, Dale Lackeyram, Todd Rideout, Yingxin Gao, Cornelis F, M, de Lange, Roger R, Hacker, y Willem C, Sauer, 2001, «Novel Methodology Allows Simultaneous Measurement of True Phosphorus Digestibility and the Gastrointestinal Endogenous Phosphorus Outputs in Studies with Pigs», The Journal of Nutrition 131 (9): 2388-96, https://doi.org/10.1093/jn/131.9,2388. 10. Just, Franziska, M, Oster, K, Büsing, Luisa Borgelt, E, Muráni, S, Ponsuksili, Petra Wolf, y K, Wimmers, 2018, «Lowered dietary phosphorus affects intestinal and renal gene expression to maintain mineral homeostasis with immunomodulatory implications in weaned piglets», BMC Genomics 19, https://doi.org/10.1186/s12864-018-4584-2. 11. Lautrou, M,, C, Pomar, J,-Y, Dourmad, A, Narcy, P, Schmidely, y M, P, Létourneau-Montminy, 2020, «Phosphorus and Calcium Requirements for Bone Mineralisation of Growing Pigs Predicted by Mechanistic Modelling», Animal 14 (S2): s313-22, https://doi.org/10.1017/S1751731120001627. 12. Lautrou, Marion, Agnès Narcy, Jean-Yves Dourmad, Candido Pomar, Philippe Schmidely, y Marie-Pierre Létourneau Montminy, 2021, «Dietary Phosphorus and Calcium Utilization in Growing Pigs: Requirements and Improvements», Frontiers in Veterinary Science 8 (noviembre):734365, https://doi.org/10,3389/ 13.Leske, K,, y C, Coon, 2002, «The Development of Feedstuff Retainable Phosphorus Values for Broilers», Poultry Science 81 (11): 1681-93, https://doi.org/10.1093/ps/81,11,1681. 14. Liu, J, B,, X, Y, Shen, H, X, Zhai, L, Chen, y H, F, Zhang, 2017, «Dietary sources of phosphorus affect postileal phosphorus digestion in growing pigs1», Journal of Animal Science 95 (10): 4490-98, https://doi.org/10.2527/jas2017.1426. 15. Liu, J,, D, W, Bollinger, D, R, Ledoux, y T, L, Venum, 2000, «Effects of dietary calcium:phosphorus ratios on apparent absorption of calcium and phosphorus in the small intestine, cecum, and colon of pigs», Journal of Animal Science 78 (1): 106-9, https://doi.org/10.2527/2000.781106x. 16. LR – Animal Nutrition, WIAS, P, Bikker, y M,C, Blok, 2017, «Phosphorus and Calcium Requirements of Growing Pigs and Sows», Wageningen: Wageningen Livestock Research, https://doi.org/10.18174/424780. 17. Nath, S, 2009, «Energy transfer from adenosine triphosphate: quantitative analysis and mechanistic insights,» The journal of physical chemistry, B 113 5:1533-37, https://doi.org/10.1021/jp809678n. 18. National Research Council, 2012, Nutrient requirements of swine, 11th edition, 2012, Vol, 1, Washington, D,C,: National Academies Press, https://doi.org/10.17226/13298. 19. Petersen, G, I,, C, Pedersen, M, D, Lindemann, y H, H, Stein, 2011, «Relative Bioavailability of Phosphorus in Inorganic Phosphorus Sources Fed to Growing Pigs», Journal of Animal Science 89 (2): 460-66, https://doi.org/10.2527/jas.2009-2161. 20. Poulsen, H, D, A, W Jongbloed, P Latimier, y J, A Fernández, 1999, «Phosphorus consumption, utilisation and losses in pig production in France, The Netherlands and Denmark», Livestock Production Science 58 (3): 251-59, https://doi.org/10.1016/S0301-6226(99)00013-5. 21. Rodehutscord, M,, M, Faust, y E, Pfeffer, 1999, «The course of phosphorus excretion in growing pigs fed continuously increasing phosphorus concentrations after a phosphorus depletion», Archiv für Tierernaehrung 52 (4): 323-34, https://doi.org/10.1080/17450399909386171. 22. Selle, Peter H,, y Velmurugu Ravindran, 2008, «Phytate-Degrading Enzymes in Pig Nutrition», Livestock Science 113 (2-3): 99-122, https://doi.org/10.1016/j.livsci.2007.05.014. 23. She, Yue, Defa Li, y Shuai Zhang, 2017, «Methodological Aspects of Determining Phosphorus Digestibility in Swine: A Review», Animal Nutrition (Zhongguo Xu Mu Shou Yi Xue Hui) 3 (2): 97-102, https://doi.org/10.1016/j.aninu.2017.02.003. 24. She, Yue, Qiuyun Wang, Hans H, Stein, Ling Liu, Defa Li, y Shuai Zhang, 2018, «Additivity of Values for Phosphorus Digestibility in Corn, Soybean Meal, and Canola Meal in Diets Fed to Growing Pigs», Asian-Australasian Journal of Animal Sciences 31 (8): 1301-7, https://doi.org/10.5713/ajas.17.0547. 25. Shen, Yingran, Ming Z, Fan, Ayodele Ajakaiye, y Tania Archbold, 2002, «Use of the Regression Analysis Technique to Determine the True Phosphorus Digestibility and the Endogenous Phosphorus Output Associated with Corn in Growing Pigs», The Journal of Nutrition 132 (6): 1199-1206, https://doi.org/10.1093/jn/132.6.1199. 26. Silva, V, M,, F, Putti, P, White, y A, R, Reis, 2021, «Phytic acid accumulation in plants: Biosynthesis pathway regulation and role in human diet,» Plant physiology and biochemistry: PPB 164:132-46, https://doi.org/10.1016/j.plaphy.2021.04.035. 27. Stein, H, H,, B, Seve, M, F, Fuller, P, J, Moughan, y C, F, M, de Lange, 2007, «Invited Review: Amino Acid Bioavailability and Digestibility in Pig 28. Suttle, N, F, 2010, Mineral Nutrition of Livestock, 4th ed, Wallingford, Oxfordshire, UK ; Cambridge, MA: CABI, 29. Underwood, E,J,, K, N,, y N, F, Suttle, 2000, «The Mineral Nutrition of Livestock»,, British Journal of Nutrition 84 (3): 393-393, https://doi.org/10.1017/S0007114500001689. 30. Yang, Xiao-e, Xiang Wu, Hu-lin Hao, y Zhen-li He, 2008, «Mechanisms and Assessment of Water Eutrophication», Journal of Zhejiang University SCIENCE B 9 (3): 197-209, https://doi.org/10.1631/jzus.B0710626. 31. Zhai, Hengxiao, Olayiwola Adeola, y Jingbo Liu, 2022, «Phosphorus nutrition of growing pigs», Animal Nutrition 9 (junio):127-37, https:// doi.org/10.1016/j.aninu.2022.01.010. 32. Rutherfurd, S. M., T. K. Chung, y P. J. Moughan. 2014. «Effect of Microbial Phytase on Phytate P Degradation and Apparent Digestibility of Total P and Ca throughout the Gastrointestinal Tract of the Growing Pig1». Journal of Animal Science 92 (1): 189-97. https://doi.org/10.2527/jas.2013-6923. |
[/registrados]