

Desde que el microorganismo patógeno causante de disentería hemorrágica porcina fue descrito por primera vez en el año 1921, se ha denominado de diferentes maneras de acuerdo a su transmisión y patogenicidad. Comenzó considerándose como Treponema hyodysenteriae, para luego clasificarse en el género Serpulina. Hoy en día se conoce como Brachyspira hyodysenteriae (B. hyodysenteriae).



Las principales características de B. hyodysenteriae se describen a continuación:
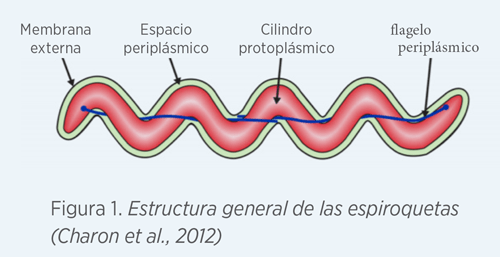
- Es una bacteria Gram –, anaerobia y con forma espiral. Posee flagelos aportándole motilidad en medios altamente viscosos. El número de flagelos es variable, de 7 a 14 (Hampson, 2006)
- Posee multitud de factores de virulencia: lipopolisacáridos (endotoxinas), hemolisinas (actividad hemolítica), actividad de quimiotaxis por la mucina, motilidad, actividad NADH oxidasa (tolerancia al oxígeno), proteínas de membrana…

Aunque la patogenia de la enfermedad se ha descrito en profundidad en los últimos años, todavía sigue habiendo preguntas que no se pueden responder con exactitud. Por ejemplo, se han establecido hipótesis para tratar de entender mejor por qué Brachyspira es capaz de colonizar la mucosa del colon durante largos periodos de tiempo sin ser eliminada del hospedador.
LA PATOGENIA, tratando de aportar algo de claridad
Aunque existen varias vías de entrada de la enfermedad, las principales fuentes de infección son los cerdos infectados de forma subclínica, los camiones contaminados que entran y salen de las granjas y los roedores, los cuales pueden eliminar en heces la bacteria durante un periodo aproximado de 6 meses.
![]()

Un factor clave para determinar si se va a desarrollar la enfermedad en los animales es el ambiente en el que se encuentra la bacteria.


Por otro lado, se ha descrito que la bacteria se detecta en heces entre 1 y 4 días antes de que comiencen los síntomas (Kinyon et al., 1977), por lo que una vez que comienza la diarrea la presión de infección ya es elevada.
A continuación, en la Figura 2 se resume la patogenia de la enfermedad:
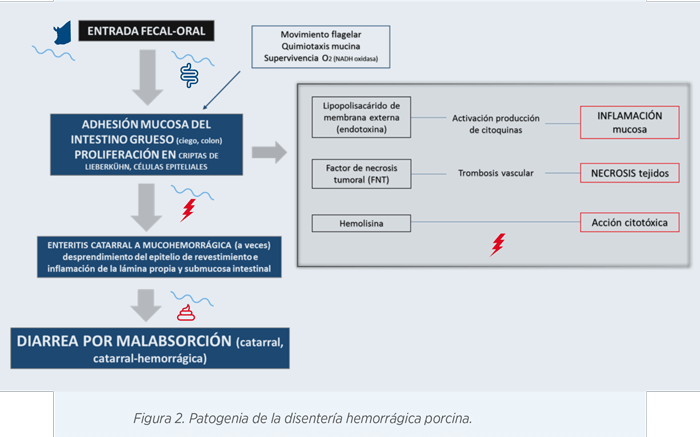



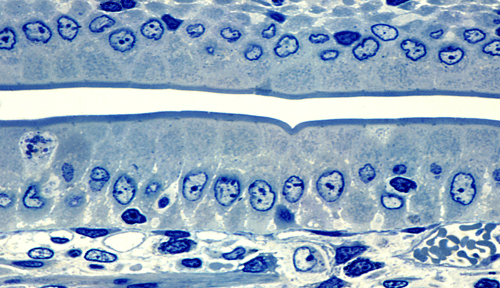
- B. hyodysenteriae coloniza primero el epitelio del ciego y colon para luego llegar a las células epiteliales y células caliciformes.
- Posteriormente, alcanza las criptas del colon las cuales son un lugar idóneo para su proliferación ya que son zonas anaeróbicas con alta motilidad, lo que permite a la bacteria desplazarse y diseminarse.
La bacteria no penetra en los enterocitos, sino que se une al epitelio mediante unos mecanismos de adhesión. Estos mecanismos podrían ser las proteínas de membrana externa (OMPs), las cuales se explican posteriormente.


El papel de las proteínas de membrana externa (OMPS)
La membrana externa (OM) de las espiroquetas está compuesta principalmente por proteínas de membrana (OMPs), porinas y receptores de proteínas. La membrana interna (IM) se encuentra adherida al peptidoglicano (pg). En el espacio periplásmico es donde se encuentran los flagelos periplásmicos (Ef).
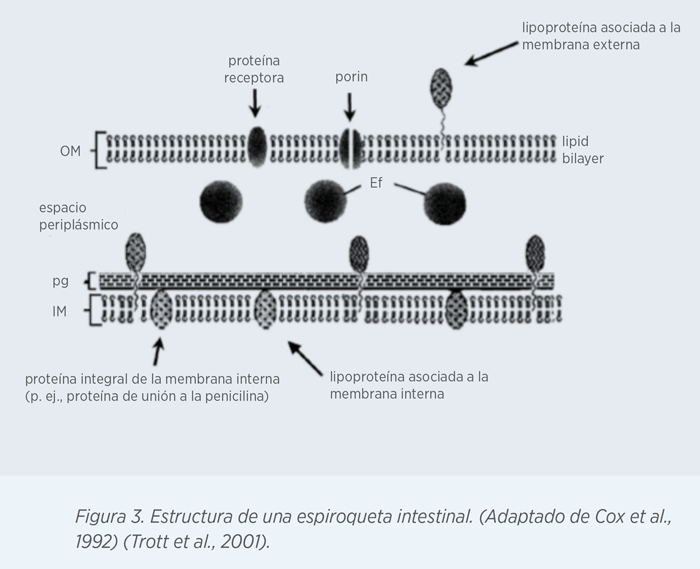

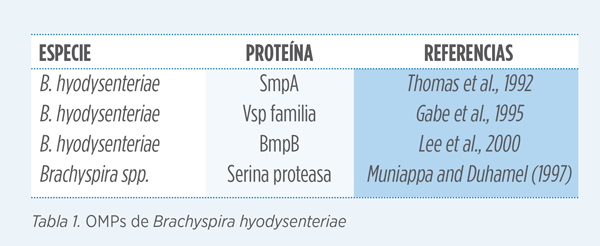
En general, son pocas las proteínas de membrana externas (OMPs) que se han caracterizado para B. hyodysenteriae. Estas proteínas se expresan en la superficie de la bacteria y se consideran mediadoras en la adhesión a las células del hospedador.
Podrían representar un papel importante en la colonización y el desarrollo de la enfermedad. Estas OMPs se unirían a los componentes de la matriz extracelular del epitelio intestinal sin llegar a penetrar en los enterocitos (Cullen et al., 2004). Una vez adherida a la superficie del epitelio, la bacteria pone en marcha todo su arsenal de patogenicidad y la superficie de la mucosa acaba siendo erosionada desencadenando los síntomas y lesiones previamente descritos.

Por otro lado, las OMPs podrían ser los principales objetivos de la respuesta inmune del hospedador.
Se han establecido hipótesis para tratar de entender mejor por qué Brachyspira es capaz de colonizar la mucosa del colon durante largos periodos de tiempo sin ser eliminada del hospedador.
Para ello, las OMPs podrían disminuir su actividad durante las fases agudas de la disentería, lo que correspondería con un mecanismo de evasión de la respuesta inmune y el consecuente establecimiento de una infección crónica.
Las OMPs también pueden regularse en función del estado del ambiente, por ejemplo, es posible que actúen directamente sobre el hospedador en respuesta a un estímulo ambiental como la baja concentración de hierro (Li et al., 1995).
Se han hallado diferencias en la membrana externa de Brachyspira con respecto al resto de espiroquetas. Por ejemplo, la presencia de colesterol en la misma no se ha reportado en otros procariotas.
Este colesterol puede conferir unas características únicas a la membrana, como, por ejemplo, una reducción de la densidad (Trott et al., 2001).

HERRAMIENTAS DE CONTROL
Se puede afirmar que en España la incidencia de granjas que cursan con diarrea hemorrágica ha aumentado en los últimos meses debido principalmente al uso responsable de antibióticos en producción animal, lo que ha llevado a una disminución progresiva de varias categorías de antibióticos.


Para aquellos casos en los que se tenga que llevar a cabo un tratamiento antibiótico, se hace indispensable la determinación de la concentración mínima inhibitoria para molécula antibiótica.
En base a este diagnóstico, se deberá elegir aquella molécula que mejor resultado haya obtenido. Este protocolo de actuación ha favorecido una mejor utilización de los antibióticos y, por consiguiente, una reducción de la resistencia a los mismos.
Existen otros factores que contribuyen a esta mayor prevalencia de disentería como:
- El aumento del número de granjas
- El tamaño de las mismas
- La existencia de zonas geográficas que actualmente abarcan una alta densidad de animales
Todo esto genera más desplazamientos de camiones con y/o sin cerdos.

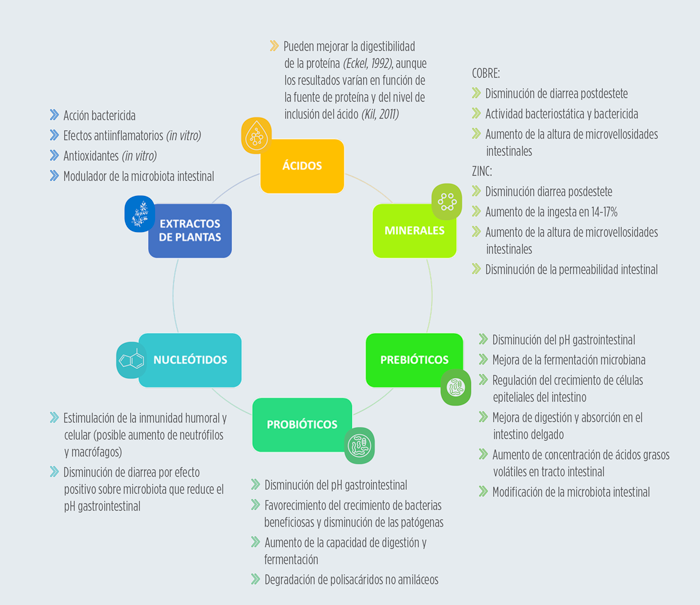
Hoy en día existen múltiples estrategias de actuación frente a la disentería porcina, aparte de la antibioterapia. La eficacia de estos productos es variable, debido en parte a que el mecanismo de acción de B. hyodysenteriae es complejo.
A continuación, se resumen los principales productos considerados como aditivos que se están implementando para el control de los problemas digestivos.

CONCLUSIONES
Para llevar a cabo las medidas de control de la disentería porcina se hace necesario el entendimiento en la medida de lo posible de cómo B. hyodysenteriae actúa en el organismo de los hospedadores.
Para ello, se están llevando a cabo numerosos estudios que determinan en qué momento y en qué parte de la bacteria se debería actuar.
Teniendo en cuenta la densidad ganadera de algunas zonas de España, la erradicación de la disentería porcina se plantea solo en aquellas zonas donde existen unos mínimos requisitos para obtener resultados positivos y duraderos en el tiempo.


Este control comienza por la bioseguridad y continúa con el resto de medidas de control que variarán de un caso a otro, desde el uso de antibióticos hasta la utilización de alternativas no medicamentosas ya sea en forma de minerales, prebióticos, probióticos etc.
BIBLIOGRAFÍA
1. Bellgard, M.I.; Wanchanthuek, P.; La, T.; Ryan, K.; Moolhuijzen, P.; Albertyn, Z.; Shaban, B.; Motro, Y.; Dunn, D.S.; Schibeci, D., 2009. Genome sequence of the pathogenic intestinal spirochaete Brachyspira hyodysenteriae reveals adaptations to its lifestyle in the porcine large intestine. PLoS One 2009, 4, e4641, doi:10.1371/journal.pone.0004641
2. Burrough E.R. 2017. Swine Dysentery: Etiopathogenesis and Diagnosis of a Reemerging Disease. Veterinary Pathology, Vol. 54(1) 22-31
3. Charon N.W., Cockburn A., Li C., Liu J., Miller K.A., Miller M.R., Motaleb M., and Wolgemuth C.W. 2012. The unique paradigm of spirochete motility and chemotaxis. Annu Rev Microbiol.; 66: 349–370.
4. Cox D.L., Chang P., Mcdowall A.W., Radolf J.D. 1992. The Outer Membrane, Not a Coat of Host Proteins, Limits Antigenicity of Virulent Treponema pallidum. Infection and immunity, p. 1076-1083
5. Cullen P, Haake D, Adler B. Outer membrane proteins of pathogenic spirochetes. FEMS Microbiol Rev. 2004 Jun;28(3):291-318
6. Eckel B, Kirchgessner M, Roth FX. Influence of formic acid on daily weight gain, feed intake, feed conversion rate and digestibility. The nutritive value of organic acids in the rearing of piglets. J Anim Physiol Anim Nutr 1992;62:93e100.
7. Gabe J.D., Chang R.J., Slomiany R., Andrews W.H. and McCaman M.T. (1995). Isolation of extracytoplasmic proteins from Serpulina hyodysenteriae B204 and molecular cloning of the fla B1 gene encoding a 38-kilodalton flagella protein. Infection and Immunity 63: 142–148.
8. Hampson, D.J.; Fellstrom, C.; Thomson, J.R. Swine dysentery. In Diseases of Swine; Straw, B.E., Zimmerman, J.J., D’Allaire, S., Eds.; Blackwell Publishing Professional: Ames, IA, USA, 2006; pp. 785–805.
9. Kil DY, Kwon WB, Kim BG. Dietary acidifiers in weanling pig diets: a review. Rev Colombiana Ciencias Pecuarias 2011;24:231e47.
10. Kynion J.M., Harris D.L., GLOCK R.D. 1977. Enteropathogenicity of Various Isolates of Treponema hyodysenteriae. Infection and immunity; p. 638-646
11. Lee B.J., La T., Mikosza A.S.J. and Hampson D.J. (2000). Identification of the gene encoding BmpB, a 30 kDa outer envelope lipoprotein of Brachyspira (Serpulina) hyodysenteriae and immunogenicity of recombinant BmpB in mice and pigs. Veterinary Microbiology 76: 245–257.
12. Li Z., Foiry B., Jacques M. 1995. Growth of Serpulina (Treponema) hyodysenteriae under Iron-restricted Conditions. Can J Vet Res 1995; 59: 149-153
13. Liu Y., Espinosa C.D., Abelilla J.J., Casas G.A., Lagos L.V., Lee S.A., Kwon W.B., Mathai J.K., Navarro D., Jaworski N.W., Stein H.H. 2018. Non-antibiotic feed additives in diets for pigs: A review. Journal of Animal nutrition
14. Muniappa N., Duhamel G.E. 1997. Outer membrane-associated serine protease of intestinal spirochetes. FEMS Microbiology Letters, Volume 154, Issue 1, Pages 159–164
15. Thomas W., Sellwood R. 1992. Monoclonal antibodies to a 16-kDa antigen of Serpulina (Treponema) hyodysenteriae. J. Med. Microbiol. – Vol. 37 (1992), 214-220
16. Trott D.J.,. Alt D.P., Zuerner1 R.L., Wannemuehler M.J., Stanton T.B. 2001. The search for Brachyspira outer membrane proteins that interact with the host. Animal Health Research Reviews 2(1); 19–30