


En las personas, el concepto de subfertilidad varía con respecto a la producción animal. La subfertilidad humana responde a una ausencia de concepción durante un tiempo prolongado, mientras que el término infertilidad se utiliza como sinónimo de esterilidad, con la aparición únicamente de gestaciones esporádicas.
![]()



![]()

¿Qué es un verraco subfértil y cuál es su incidencia? 



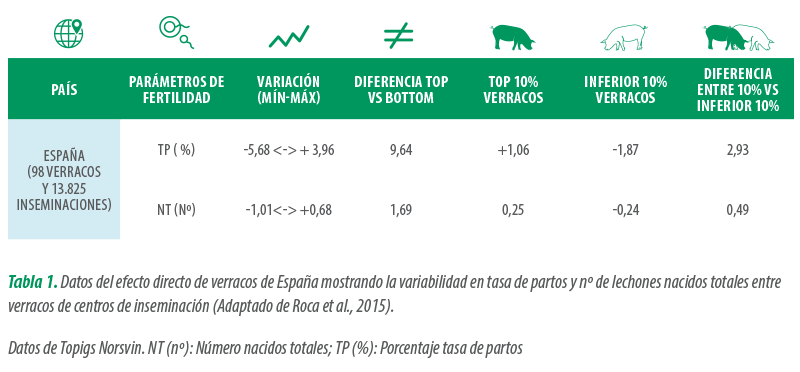
Se trata de un aspecto difícil de determinar ya que, como algunos autores indican, la variabilidad de fertilidad explicada entre granjas por los parámetros relacionados con el verraco únicamente representa el 6% de la fertilidad in vivo (Broekhuijse et al., 2012).
La estimación de algunos autores sobre la incidencia de verracos subfértiles es alrededor del 5% (I. Parrilla, 2016), aunque se trata de un porcentaje difícil de establecer.
En la Tabla 1, se muestran datos de [registrados]la variabilidad en cuanto a la tasa de partos y la prolificidad de 98 verracos.



Defectos compensables y no compensables
Algunos de los defectos que podrían causar una baja tasa de partos y/o prolificidad, como una reducción en la motilidad o un elevado porcentaje de formas anormales, pueden verse enmascarados o compensados por la cantidad de espermatozoides que se utilizan en las dosis seminales hoy en día. Sin embargo, hay una serie de atributos o defectos que no se pueden compensar.
Estos defectos no imposibilitan que el espermatozoide entre en contacto con el ovocito y provoque el bloqueo espermático, de manera que, aunque se añada una mayor cantidad de espermatozoides, no se evitará la afectación de los parámetros reproductivos (A. Bolarín, 2018).
Estas características se categorizan en defectos compensables y no compensables (Braundmeier & Miller, 2001).
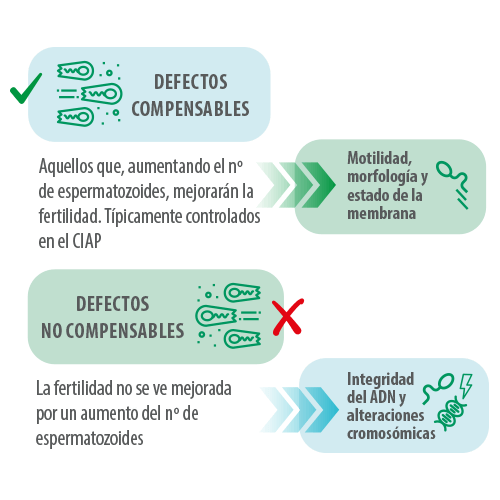

La presencia de estos defectos no compensables serán los que caractericen a los verracos subfértiles, aunque también debemos tener en cuenta que los defectos compensables, son “compensables” hasta cierto punto, y dependerán del grado de afectación o de la posible suma de varios factores.
![]()

¿Cuáles son las causas más frecuentes de subfertilidad y sus consecuencias?

![]()




![]()




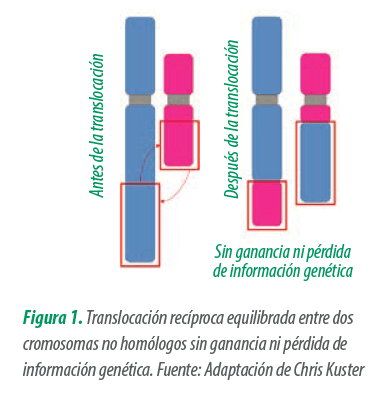

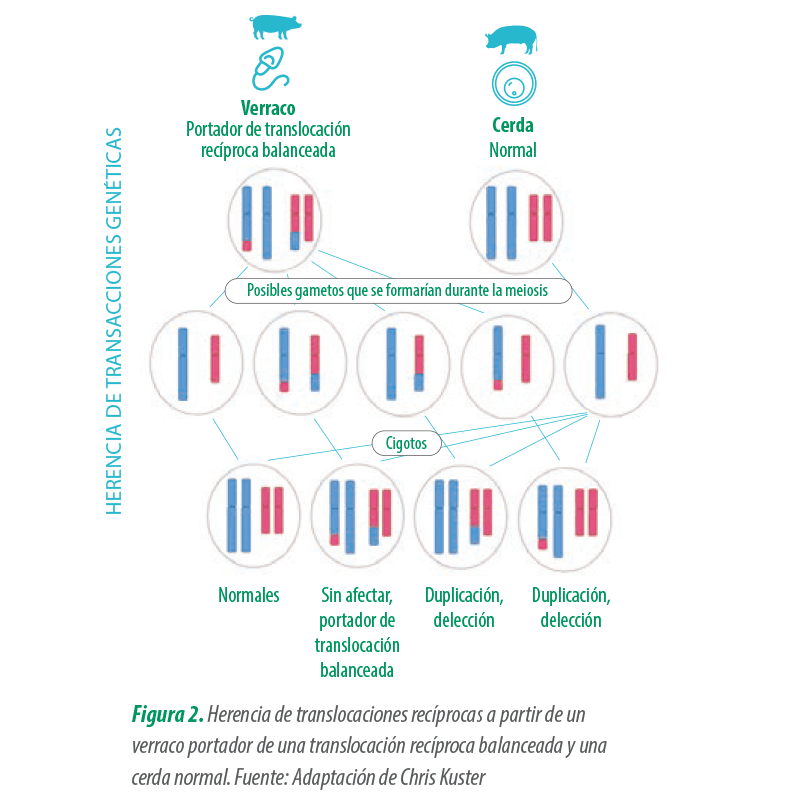






Las translocaciones pueden aparecer de forma espontánea o de novo, debido a una rotura cromosómica y un fallo en la reorganización durante la formación de los gametos (meiosis), con una incidencia de entre el 0,5% y 0,7% (datos de Kuster Research Center).
Este tipo de alteración posee una fuerte correlación con verracos diagnosticados como hipoprolíficos, con un 50% en Europa (A. Ducos et al., 2008) y un 48,4% en Estados Unidos (Kuster Research Center).

![]()


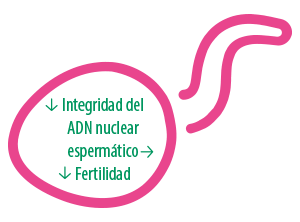
La otra principal problemática de los defectos no compensables es la fragmentación del ADN nuclear espermático (DFI, DNA fragmentation index).
![]()

La evaluación de la integridad del ADN nos permite identificar roturas del material genético del espermatozoide, de modo que cuanto mayor sea el daño, menor será la integridad del ADN por lo que incrementará la probabilidad de que disminuya su fertilidad.

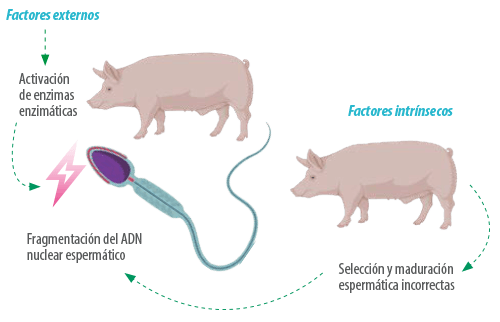
A diferencia del parámetro anterior, donde la aparición de la problemática se debía a una generación de novo o por la aparición de portadores, el daño en la integridad del ADN puede deberse a factores externos que pueden causar la activación de enzimas espermáticas que induzcan el daño en el ADN o a factores intrínsecos del verraco, que deriven en una selección y maduración espermática incorrectas, debidas a componentes genéticos del verraco no conocidos.

La evaluación de la fragmentación del ADN por múltiples técnicas, como el test SCSA y TUNEL (Waberski et al., 2011), y el estudio citogenético para la detección de alteraciones cromosómicas mediante la técnica del cariotipado (Sánchez-Sánchez et al., 2019), permiten la detección de machos subfértiles debidos a estas problemáticas.
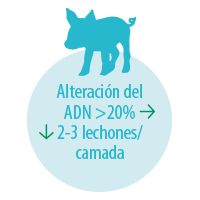
El porcentaje de daño del ADN nuclear espermático en el verraco es generalmente bajo (<5%) y se caracteriza por no verse incrementado con la conservación de las dosis seminales (Waberski et al., 2011).
Al tratarse de un parámetro que puede verse alterado por factores externos, hace que no se conozcan, o no se puedan dar de forma fiable, estimaciones del porcentaje de animales afectados por esta problemática.


Al igual que en el caso de las translocaciones recíprocas, la consecuencia de una alteración de la integridad del ADN nuclear espermático será una reducción del tamaño de camada debido al fallo en el desarrollo embrionario.


Todavía hay múltiples líneas de investigación que en un futuro permitirán conocer o predecir la capacidad fecundante de un verraco con mayor exactitud. Por ello, es más que probable que en los próximos años se desarrollen herramientas y tests que permitan llevar a cabo un panel de evaluación de diversos parámetros, que nos permitan estimar de una forma más exacta la capacidad reproductiva de los verracos.
BIBLIOGRAFÍA
Braundmeier, A. G., & Miller, D. J. (2001). The search is on: Finding accurate molecular markers of male fertility. Journal of Dairy Science, 84(9), 1915–1925. https://doi.org/10.3168/jds.S0022-0302(01)74633-4
Broekhuijse, M. L. W. J., Feitsma, H., & Gadella, B. M. (2012). Artificial insemination in pigs: Predicting male fertility. Veterinary Quarterly, 32(3–4), 151–157. https://doi.org/10.1080/01652176.2012.735126
Danielak-Czech, B., Świtoński, M., & SŁota, E. (1997). First identification of reciprocal translocations in Polish pigs. Journal of Animal Breeding and Genetics, 114(1), 69–78. https://doi.org/10.1111/j.1439-0388.1997.tb00493.x
Ducos, A., Revay, T., Kovacs, A., Hidas, A., Pinton, A., Bonnet-Garnier, A., Molteni, L., Slota, E., Switonski, M., Arruga, M. V., Van Haeringen, W. A., Nicolae, I., Chaves, R., Guedes-Pinto, H., Andersson, M., & Iannuzzi, L. (2008). Cytogenetic screening of livestock populations in Europe: An overview. Cytogenetic and Genome Research, 120(1–2), 26–41. https://doi.org/10.1159/000118738
Ducos, Alain, Berland, H. M., Bonnet, N., Calgaro, A., Billoux, S., Mary, N., Garnier-Bonnet, A., Darré, R., & Pinton, A. (2007). Chromosomal control of pig populations in France: 2002-2006 Survey. Genetics Selection Evolution, 39(5), 583–597. https://doi.org/10.1051/gse:2007023
Foxcroft, G. R., Dyck, M. K., Ruiz-Sanchez, A., Novak, S., & Dixon, W. T. (2008). Identifying useable semen. Theriogenology, 70(8), 1324–1336. https://doi.org/10.1016/j.theriogenology.2008.07.015
Gnoth, C., Godehardt, E., Frank-Herrmann, P., Friol, K., Tigges, J., & Freundl, G. (2005). Definition and prevalence of subfertility and infertility. Human Reproduction, 20(5), 1144–1147. https://doi.org/10.1093/humrep/deh870
Quach, A. T., Revay, T., Villagomez, D. A. F., Macedo, M. P., Sullivan, A., Maignel, L., Wyss, S., Sullivan, B., & King, W. A. (2016). Prevalence and consequences of chromosomal abnormalities in Canadian commercial swine herds. Genetics Selection Evolution, 48(1), 1–7. https://doi.org/10.1186/s12711-016-0246-5
Roca, J., Broekhuijse, M. L. W. J., Parrilla, I., Rodriguez-Martinez, H., Martinez, E. A., & Bolarin, A. (2015). Boar Differences In Artificial Insemination Outcomes: Can They Be Minimized? Reproduction in Domestic Animals = Zuchthygiene, 50, 48–55. https://doi.org/10.1111/rda.12530
Sánchez-Sánchez, R., Gómez-Fidalgo, E., Pérez-Garnelo, S., Martín-Lluch, M., & De la Cruz-Vigo, P. (2019). Prevalence of chromosomal aberrations in breeding pigs in Spain. Reproduction in Domestic Animals, 54(S4), 98–101. https://doi.org/10.1111/rda.13540
Silva, P. F. N., & Gadella, B. M. (2006). Detection of damage in mammalian sperm cells. Theriogenology, 65(5 SPEC. ISS.), 958–978. https://doi.org/10.1016/j.theriogenology.2005.09.010
Waberski, D., Schapmann, E., Henning, H., Riesenbeck, A., & Brandt, H. (2011). Sperm chromatin structural integrity in normospermic boars is not related to semen storage and fertility after routine AI. Theriogenology, 75(2), 337–345. https://doi.org/10.1016/j.theriogenology.2010.09.004
Inmaculada Parrilla. Biomarcadores de fertilidad en el semen de verraco. Revista Anaporc, nº 135.
Alfonso Bolarín. Fragmentación del ADN nuclear espermático como causa de subfertilidad indetectable en un verraco de un centro de inseminación. Revista Anaporc, nº 152.

[/registrados]