En el año 2009 emergió una variante del subtipo H1N1 de la Influenza A que afectaba a la especie humana, escapando a la inmunidad existente. Los primeros casos se dieron en México, pero el virus se propagó rápidamente por todo el Mundo.
Esta situación llevó a que la OMS declarara la fase 6 de alerta de pandemia (epidemia a nivel global), lo que en su momento generó una gran alarma.
Sin embargo, las predicciones iniciales sobre la virulencia de este nuevo virus se sobreestimaron y la mortalidad que generó a nivel de la especie humana fue incluso menor que la que provoca la gripe estacional (alrededor de medio millón de personas al año).
Posteriormente se descubrió que este nuevo linaje del H1N1, que se denominó H1N1 pandémico (panH1N1), se había generado en la especie porcina tras varias décadas de evolución y reordenaciones entre virus de origen humano, aviar y porcino.
Un año después de su aparición, la OMS decretó que la fase de alerta por pandemia había concluido. Sin embargo el panH1N1 ha continuado circulando en la especie humana, hasta el punto de que las vacunas actuales incorporan actualizaciones de este virus.
La especie porcina jugó un papel determinante en la aparición del panH1N1, pero fue la especie humana la que se encargó de transmitirlo a nivel global.
 De forma retrógrada, el panH1N1 ha acabado infectando a muchas de las explotaciones de porcino como consecuencia del contacto con la especie humana. El panH1N1 se detecta en la mayoría de continentes e incluso en países (como Noruega o Australia) que eran libres de Influenza en porcino. En Europa el panH1N1 ya se detecta en todos los países con producción porcina importante.
De forma retrógrada, el panH1N1 ha acabado infectando a muchas de las explotaciones de porcino como consecuencia del contacto con la especie humana. El panH1N1 se detecta en la mayoría de continentes e incluso en países (como Noruega o Australia) que eran libres de Influenza en porcino. En Europa el panH1N1 ya se detecta en todos los países con producción porcina importante.
¿CÓMO SE GENERÓ EL PANH1N1?
 DIVERSIDAD Y CAPACIDAD DE EVOLUCIÓN DEL VIRUS DE LA INFLUENZA PORCINA
DIVERSIDAD Y CAPACIDAD DE EVOLUCIÓN DEL VIRUS DE LA INFLUENZA PORCINA
El virus de la Influenza Porcina (SIV, de Swine Influenza Virus) tiene una gran diversidad a nivel mundial. Esto es debido a básicamente 4 factores:
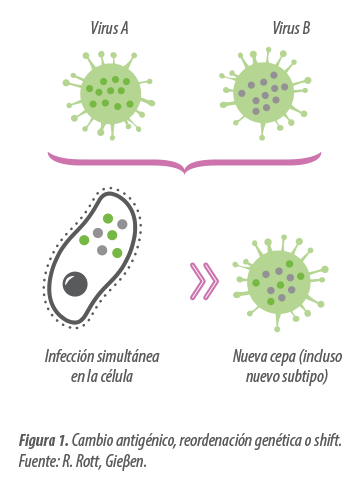
- Son virus formados por una cadena simple de ARN (igual que el PRRS), lo que conlleva que sufra alteraciones frecuentes al transcribirse (lo que se denomina deriva genética o “genetic Drift”).
- Su genoma está fragmentado en 8 segmentos independientes con capacidad de intercambiarse entre sí cuando 2 virus distintos infectan al mismo tiempo a una célula (lo que se denomina reordenamiento genético o “genetic Shift”).
- El porcino tiene la capacidad de infectarse con virus procedentes de otras especies, en especial de la especie humana y de forma más ocasional, de la aviar. Esto le permite incorporar virus de origen humano o aviar, adaptarlos y reorganizarlos con el potencial de, posteriormente, transmitírselos a la especie humana.
- La segregación geográfica de las distintas grandes zonas de producción porcina hace que linajes de SIV inicialmente parecidos se vayan diferenciando con el tiempo al evolucionar de forma separada, resultando en virus significativamente distintos entre sí. Los movimientos de cerdos entre continentes pueden, de forma ocasional, trasladar estas cepas a lugares muy distantes.
El panH1N1 se generó como fruto de estos 4 puntos comentados y tras mucho tiempo de evolución. Para comprender cómo se generó, resulta imprescindible describir mejor el fenómeno de reordenación genética y la evolución del virus, especialmente en Europa y Norte América.
 REORDENAMIENTO GENÉTICO (GENETIC SHIFT)
REORDENAMIENTO GENÉTICO (GENETIC SHIFT)
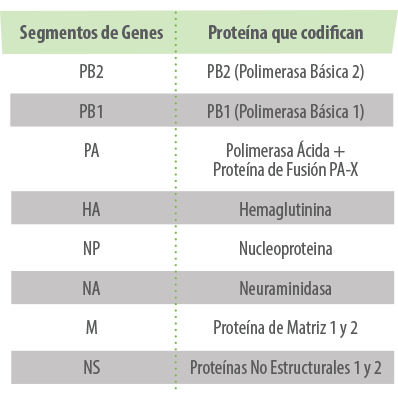
Los 8 segmentos del genoma codifican 11 proteínas, 2 externas (la Hemaglutinina (HA) y Neuraminidasa (NA)) y 9 internas.
HEMAGLUTININA & NEURAMINIDASA
La HA es la responsable de la unión del virus a la célula y la NA a su salida, por lo que son las máximas responsables de la infección y la respuesta inmunitaria. Según estas dos proteínas, los virus de la Influenza A se clasifican en diferentes subtipos.
En el caso del porcino, podemos encontrar H1N1, H1N2 y H3N2. Sin embargo, estos 3 subtipos tienen algunos matices muy importantes que se deben explicar con mayor profundidad:
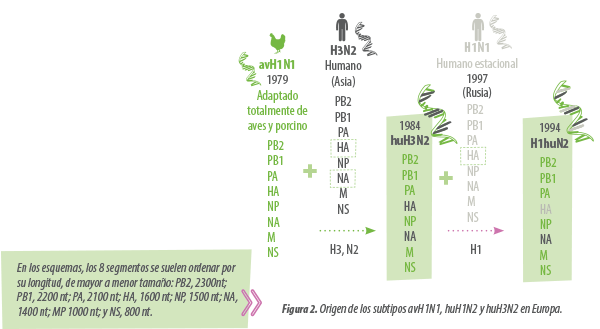
El H1N1 que circula actualmente en Europa y Asia es de procedencia enteramente aviar (sus 8 segmentos). Este virus, que denominaremos avH1N1, se adaptó completamente a la especie porcina en 1979, reemplazando al anterior H1N1 clásico (csH1N1) que era una evolución del virus que generó la pandemia de 1918 (gripe Española).
El H3N2 se adaptó al porcino en 1984, cuando el H3N2 humano estacional se reordenó con el avH1N1. Mantuvo los 6 segmentos que codifican para las proteínas internas del avH1N1 e incorporó la HA y la NA del H3N2 humano. Debido al origen de su HA y NA, se denomina huH3N2.
El H1huN2 se generó en 1994 al reordenarse la HA de un H1N1 humano de 1977 (gripe rusa) con el huH3N2 que ya circulaba en la cabaña porcina. El huH1N2 resultante no muestra buena protección cruzada con el avH1N1, porque a pesar de que ambas HA son de tipo H1, se parecen poco porque una es de procedencia humana y la otra aviar.
El H1N1 es una evolución del csH1N1 (gripe Española), por lo que no tienen nada que ver con el avH1N1 Euroasiático.
El H3N2 ya se detectaba en porcino a finales de los ‘70, pero se trataba de un virus cercano al H3N2 estacional humano, sin una excesiva virulencia. Sin embargo, a finales de los ’90 se sucedieron una serie de reordenaciones que aumentaron la diversidad del SIV en Norteamérica:
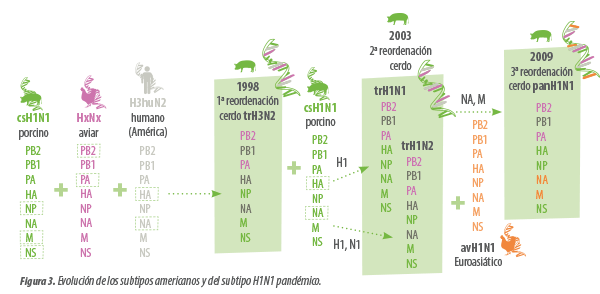
1.- En 1998 este H3N2 estacional se reordenó con:
- Un csH1N1 que aportó algunos de sus genes internos (NP/M/NS)
- Un virus aviar (HxNx) que aportó otros genes internos (PB2/PA)
El resultado fue un H3N2 más virulento, que se denomina trH3N2 (la “tr” se refiere a la triple reordenación que lo generó).
2.- Alrededor del 2003, este nuevo trH3N2 se reordenó otra vez con un csH1N1:
- Incorporando su HA para dar trH1N2
- Incorporando su HA y su NA para dar trH1N1
APARICIÓN DEL LINAJE PANH1N1
La base del panH1N1, se generó durante muchos años en la cabaña porcina norteamericana, con estas reordenaciones que culminan con la aparición del trH1N2 y trH1N1.
El paso final se dio cuando o bien el trH1N2 o bien el trH1N1 se reordenó con un avH1N1 de origen Euroasiático, que aportó los genes de la NA y M.
Según parece, esta última reordenación con el avH1N1 Euroasiático pudo haberse dado mucho antes del 2009, porque se ha comprobado que los 8 segmentos del panH1N1 llevaban establecidos en esta población como mínimo 10 años y que estaban distribuidos en un área geográfica muy extensa.
Este hecho, junto con la capacidad del porcino de adaptar virus de origen aviar y humano, ha llevado a asumir que este nuevo virus se generó en el cerdo. Sin embargo, el origen de la reordenación final con el avH1N1 Euroasiático no se ha llegado a establecer con claridad.
En Asia se detectan prácticamente todos los SIV europeos sumados a los americanos, además de tener otras variantes propias. Es presumible que el movimiento de cerdos entre Eurasia y Norte América haya facilitado la mezcla de SIV de distintos territorios mundiales, facilitando las múltiples reordenaciones que han acabado generando el panH1N1.
REORDENACIONES DEL PANH1N1 CON OTROS SUBTIPOS
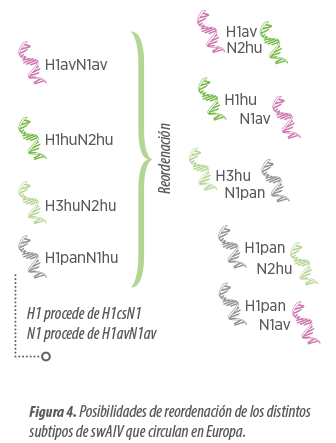
Tal y como estamos viendo, en Europa hoy en día ya podemos hablar de 4 linajes distintos de SIV:
- avH1N1
- panH1N1
- panH1N1
- huH3N2
Sin embargo, las HA y NA pueden continuar reordenándose entre sí, generando nuevas combinaciones.
En el esquema tan solo se reflejan reordenaciones de HA y NA, pero también se observan reordenaciones que contienen genes internos del virus pandémico. Estas variantes parecen tener algunas ventajas biológicas en relación a otros tipos de SIV, lo que les podría permitir ser más agresivas.
Las posibilidades de reordenación son muchas, pero hay que tener en cuenta que una vacuna que cubra los 4 subtipos circulantes de forma efectiva, cubre todas sus reordenaciones entre sí.
El SIV evoluciona de forma constante debido a la deriva genética (genetic drift) que provoca ligeros cambios, pero de forma continuada. Su capacidad de reordenarse entre sí (genetic Swift) y con genes procedentes de virus aviares y especialmente humanos, hace que puntualmente aparezcan cambios mucho más importantes con el potencial de adaptar nuevos subtipos y linajes al cerdo.
 En Europa, esto ha sucedido 4 veces en aproximadamente los últimos 40 años. Cuando esto sucede, las vacunas comerciales de SIV deben adaptarse para continuar ofreciendo una protección lo más eficaz posible.
En Europa, esto ha sucedido 4 veces en aproximadamente los últimos 40 años. Cuando esto sucede, las vacunas comerciales de SIV deben adaptarse para continuar ofreciendo una protección lo más eficaz posible.
Otro riesgo aún mayor ocurre cuando estos nuevos subtipos o linajes adaptados al cerdo resultan en virus con alto potencial patógeno, capaces de infectar a la especie humana y escapar ampliamente a la inmunidad presente. En estos casos existe el riego de generar pandemias como la que ocurrió en el año 2009.