Es frecuente encontrar varios virus respiratorios presentes de forma simultánea en infecciones en humanos y animales, incluidos los cerdos1-7. Se han descrito infecciones concurrentes de varios virus respiratorios, como el virus de la Influenza A porcina (swIAV), el virus del síndrome reproductivo y respiratorio porcino (PRRSV), el circovirus porcino de tipo 2 (PCV2) o el coronavirus respiratorio porcino (CRPV)6-8. Además, el PRRSV y el IAV, juntos o por separado, suelen ser agentes primarios o secundarios responsables del Complejo Respiratorio Porcino9,10.
Las infecciones víricas simultáneas pueden mostrar una interferencia vírica en la que un virus bloquea el crecimiento de otro11 y, dado que las infecciones mixtas del tracto respiratorio se observan con frecuencia en los cerdos, el efecto de la interacción de los patógenos en el curso de la infección merece un estudio más profundo.
El impacto de la coinfección en la gravedad y el resultado clínico no es del todo claro. Algunos estudios han demostrado que el resultado clínico de las coinfecciones víricas puede ser menos grave, o al menos no más grave, que la infección por un solo virus2,6,7,12,13. Por el contrario, otros estudios han constatado que la coinfección vírica agrava el curso clínico1,13.
Las coinfecciones de swIAV y PRRSV son comunes en el ganado porcino14-16. Ambos virus se encuentran asociados al CRP y algunos estudios indican la posibilidad de que tengan efectos sinérgicos6,17.
Debido a que muchas cepas de PRRSV tienen un potencial inmunosupresor, pueden afectar a la respuesta inmunitaria frente a otros patógenos17,18 y estudios anteriores han mostrado diversos resultados clínicos con la infección dual de PRRSV y swIAV6,7,17.
No se encontraron cambios significativos en el curso clínico de la infección en un estudio en el que los lechones fueron infectados con PRRSV y una semana después con swIAV7. En cambio, otro estudio registró una enfermedad más grave tras una coinfección en comparación con la infección única por PRRSV17. En otro estudio sobre PRRSV y swIAV, se observaron resultados clínicos variables en cerdos coinfectados con PRRSV y swIAV6.
Por lo tanto, nuestro objetivo ha sido evaluar y comparar las características clínicas, la respuesta inmunitaria y de fase aguda, la excreción viral y la carga viral de cerdos inoculados con swIAV, PRRSV o una combinación de ambos.
MATERIALESY MÉTODOS
VIRUS
El swIAV empleado en este estudio, un virus H1N1 A/Poland/Swine/14131/2014 (SwH1N1) similar al aviar, había sido aislado de un cerdo con influenza porcina aguda.
La cepa PL15-33 del PRRSV fue aislada a partir de tejido pulmonar obtenido de un cerdo con trastornos clínicos respiratorios y la secuenciación de los fragmentos ORF5 y ORF7 indicó que la cepa pertenecía al subtipo 1 del PRRSV-1.
DISEÑO EXPERIMENTAL
Se utilizaron 54 lechones de 7 semanas de edad procedentes de una granja negativa a Influenza y PRRS. Los cerdos de la explotación de origen eran seronegativos para el virus de la Enfermedad de Aujeszky y Mycoplasma hyopneumoniae. Los exámenes clínicos, serológicos y anatomopatológicos no revelaron ningún indicio de estreptococias ni de rinitis atrófica. Los lechones se asignaron aleatoriamente a 4 grupos, incluyendo un número igual de machos y hembras en cada grupo:
Grupo PRRSV (n = 14) Grupo swIAV + PRRSV (n = 14) Grupo swIAV (n = 14) Grupo control (n = 12)
Antes de comenzar el estudio, mediante PCR se comprobó que [registrados]todos los animales experimentales estaban libres de swIAV19 y PRRSV (EZ-PRRSV MPX 4.0 real-time PCR kit, Tetracore, USA), así como la ausencia de anticuerpos frente a swIAV [(pruebas de inhibición de la hemaglutinación con A/Poland/Swine/14131/2014 (H1N1), A/Poland/Swine/14131/2014 (H1N1), A/swine/ England/96 (H1N2), A/swine/Flandes/1/98 (H3N2) y H1N1 pdm-like (A/ swine/Poland/031951/12)] y frente a PRRSV (test ELISA comercial VetExpert PRRS Ab ELISA 4.0 BioNote, Corea).
El día 0, los lechones de los grupos swIAV y swIAV + PRRSV fueron inoculados por vía intranasal (IN) con SwH1N1 (107 dosis de infección de cultivo de tejidos [TCID50] en 2 mL de solución salina tamponada con fosfato (PBS). Los lechones de los grupos PRRSV y swIAV + PRRSV fueron inoculados por vía IN con PRRSV (105 TCID50 en 2 mL de PBS). Para los cerdos inoculados con ambos virus, el inóculo se mezcló justo antes de la administración IN. El volumen final de inóculo para este grupo fue el mismo (2 mL por cerdo). Por su parte, los doce cerdos inoculados con PBS (2 mL) sirvieron de control.
EXAMEN CLÍNICO
Los animales fueron examinados diariamente desde el día 7 de pre-inoculación hasta el final del periodo de estudio (día 21 día post-inoculación -dpi-) o hasta la eutanasia (2, 4 y 10 dpi). Los cerdos fueron observados y puntuados en función de los signos respiratorios:
Frecuencia respiratoria:
0 = normal
0,33 = ligeramente aumentada
0,66 = moderadamente aumentada y ligera respiración abdominal
1 = claramente aumentada y clara respiración abdominal
Secreción nasal:
0 = ausente
1 = presente
Tos:
0 = ausente
1 = presente
Estornudos:
0 = ausente
1 = presente
Anorexia:
0 = ausente
1 = presente
LAS PUNTUACIONES DETERMINADAS EN CADA CATEGORÍA SE SUMARON PARA OBTENER UNA PUNTUACIÓN CLÍNICA TOTAL PARA CADA CERDO INDIVIDUAL (0-6)
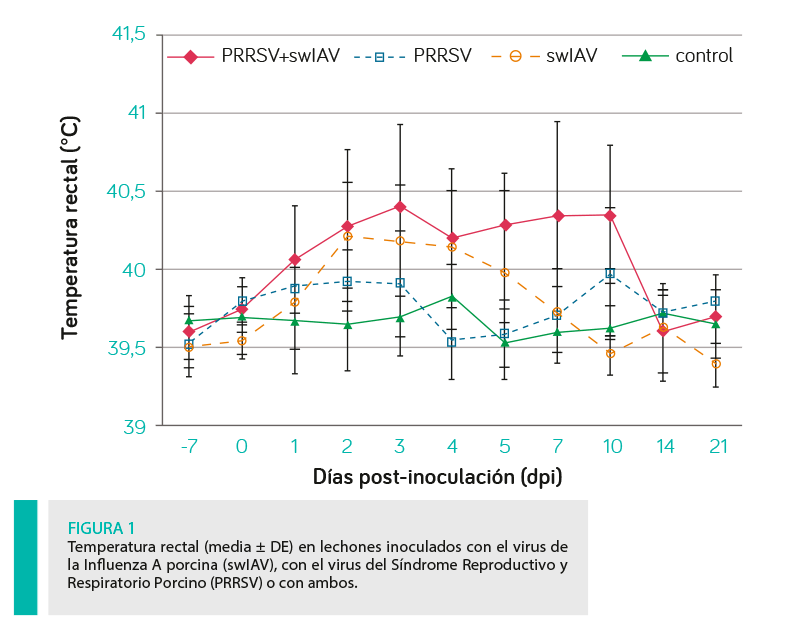
La temperatura rectal se midió diariamente, registrándose fiebre cuando la temperatura rectal alcanzaba o superaba los 40˚C. Cuando se observaba fiebre prolongada (al menos 3 días) se añadía un punto adicional a la puntuación clínica.
Se obtuvieron hisopos nasales diariamente de todos los animales y se tomaron muestras de sangre a los 7 días pre-inoculación, al día 0 (inoculación) y a los dpi 1, 2, 3, 5, 7, 10, 14 y 21. Tres lechones de los grupos inoculado y de control fueron eutanasiados a los 2, 4 y 10 dpi. El resto de los cerdos inoculados fueron eutanasiados y necropsiados a los 21 dpi.
PUNTUACIÓN PULMONAR
Las lesiones pulmonares macroscópicas se utilizaron para asignar una puntuación pulmonar20. A cada lóbulo pulmonar se le asignó un número que reflejaba un porcentaje del volumen aproximado de todo el pulmón representado:
Se asignaron 10 puntos posibles (5 para la parte dorsal y 5 para la ventral) al lóbulo anterior derecho, al lóbulo medio derecho, a la parte anterior del lóbulo anterior izquierdo y a la parte caudal del lóbulo anterior izquierdo.
Al lóbulo accesorio se le asignaron 5 puntos.
27,5 puntos (15 para la parte dorsal y 12,5 para la parte ventral) se asignaron a cada uno de los lóbulos caudales derecho e izquierdo.
La evaluación basada en este procedimiento dio como resultado una puntuación pulmonar (puntos totales = 100) correspondiente al porcentaje del pulmón afectado por neumonía
ANÁLISIS LABORATORIALES
Durante el estudio, se realizaron los siguientes análisis laboratoriales con el fin de determinar la carga viral y la respuesta inmunitaria frente a Influenza Porcina y PRRS.
Examen virológico de los hisopos y las muestras de tejido: determinación de la carga viral de swIAV en células MDCK24 y cuantificación de ARN de PRRSV mediante un kit de RT-PCR.
Serología: todos los sueros fueron sometidos a la prueba de hemaglutinación frente a swIAV y ELISA frente a PRRSV.
Prueba de proliferación de linfocitos: la prueba de proliferación de linfocitos T permitió medir las respuestas de linfocitos T específicos frente a swIAV y PRRSV de los cerdos a los 0, 7, 14 y 21 dpi21.
Medición de proteínas de fase aguda: se midieron los niveles de proteína C reactiva (CRP), haptoglobina (Hp), amiloide sérico A (SAA) y Pig-MAP (proteína de fase aguda principal en el cerdo).
RESULTADOS
EVOLUCIÓN CLÍNICA
Siete de los catorce (50%) lechones inoculados con swIAV y tres de los catorce (21,5%) inoculados con PRRSV tuvieron una fiebre persistente (>3 días) que alcanzó los 40,0-41,4˚C.
En el grupo inoculado con ambos virus, se observó fiebre persistente en diez de los catorce lechones (71,5%; Figura 1). Además, en estos lechones, la fiebre se observó durante más tiempo (hasta 10dpi) en comparación con otros grupos inoculados.
En el grupo swIAV, la fiebre alcanzó su pico máximo a los 2 dpi, mientras que en los cerdos inoculados con ambos virus se observaron 2 picos (a los 3 y 7-10 dpi), observándose una cinética de temperatura rectal similar (2 picos) en el grupo de PRRSV.
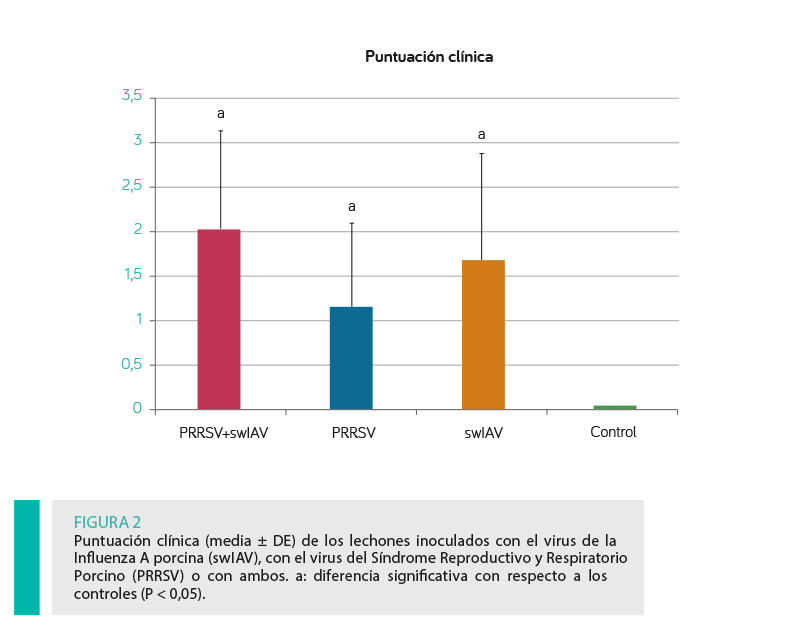
No se encontraron diferencias significativas entre las puntuaciones clínicas medias de los grupos inoculados (P ≥ 0,05; Figura 2).
Grupo swIAV + PRRSV: Trece de los catorce lechones del grupo inoculado con ambos virus presentaban al menos uno de los signos clínicos evaluados y las puntuaciones clínicas individuales en este grupo oscilaron entre 0 y 4.
Grupo swIAV: se registraron signos clínicos en once de los catorce lechones y las puntuaciones clínicas individuales oscilaron entre 0 y 3,66.
Grupo PRRSV: diez de los 14 lechones presentaron alteraciones clínicas con unas puntuaciones clínicas individuales que oscilaron entre 0 y 3.
Grupo control: no se observaron signos clínicos.
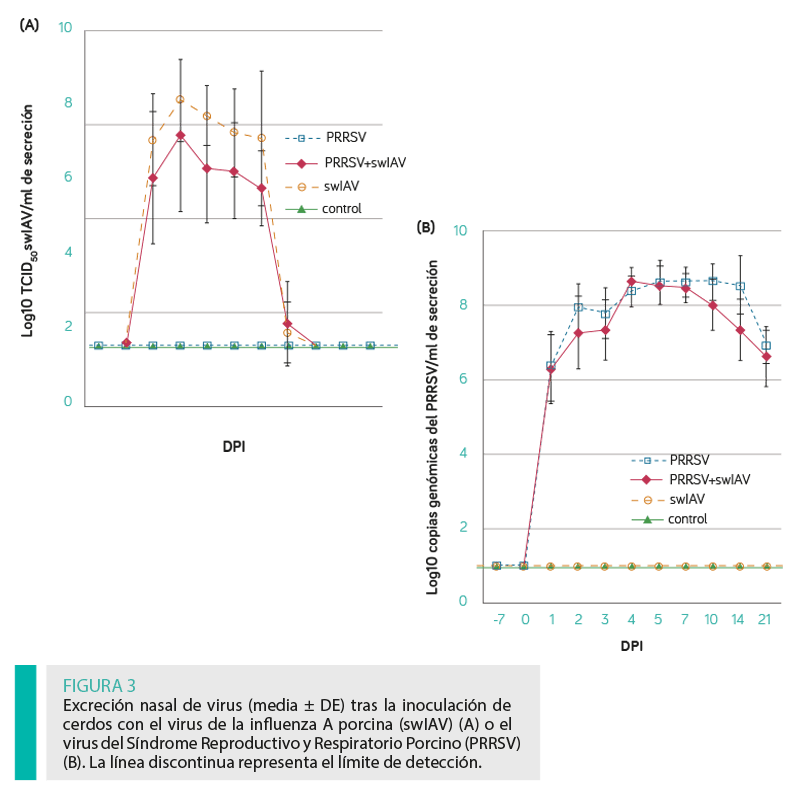
EXCRECIÓN VIRAL
El valor AUC para la excreción de swIAV y PRRSV no difirió significativamente entre los cerdos inoculados con un solo virus o con ambos (P ≥ 0,05). La dinámica de excreción de ambos patógenos (media ± DE) durante el periodo de estudio se presenta en la Figura 3A (PRRSV) y B (swIAV).
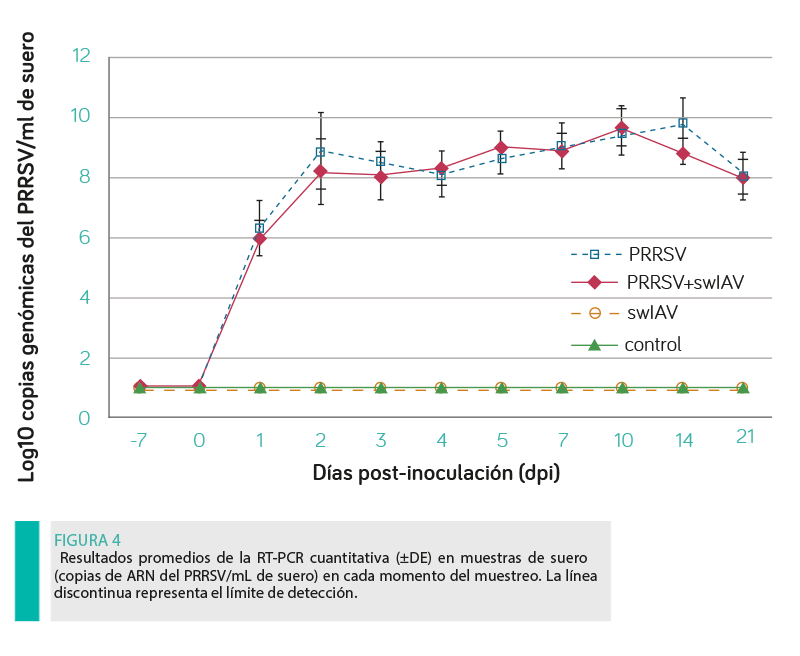
VIREMIA DE PRRSV
El AUC para la viremia de PRRSV no difirió significativamente entre los cerdos inoculados con uno de los virus o con ambos (P ≥ 0,05, Figura 4), detectándose el ARN del PRRSV en las muestras de suero de todos los animales inoculados con este virus, ya sea solo o en combinación con el swIAV.
En general, la viremia del PRRSV comenzó a partir del dpi 1 y duró hasta el final del estudio (21 dpi). No se observaron diferencias significativas entre los grupos inoculados con el PRRSV durante todo el periodo de estudio, ni se encontró ARN del PRRSV en el suero de los lechones de los grupos control y swIAV.
RESPUESTA INMUNITARIA HUMORAL
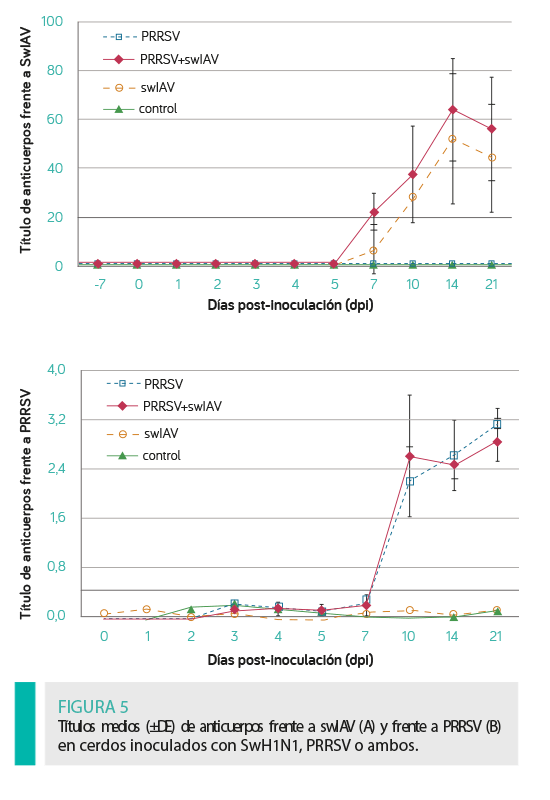
La respuesta humoral tras la inoculación de los lechones con swIAV, PRRSV o ambos se muestra en la Figura 5.
Siete de los ocho lechones del grupo inoculado con ambos virus mostraron seroconversión frente a swH1N1 a los 7 dpi.
En el grupo swIAV, solamente tres de los ocho lechones tuvieron seroconversión a los 7 dpi.
Todos los lechones inoculados con swIAV (solo o con PRRSV) mostraron seroconversión frente a este virus a los 10 DPI.
Todos los lechones inoculados con PRRSV (solo o con swIAV desarrollaron anticuerpos específicos a los 10 dpi (con la excepción de 1 cerdo en el grupo PRRSV + swIAV que seroconvirtió a los 14 dpi).
No se observaron diferencias en la magnitud de la respuesta humoral frente al swIAV y al PRRSV entre los grupos inoculados con uno de los virus o con ambos (P ≥ 0,05).
PROTEÍNAS DE FASE AGUDA
En el grupo control, las concentraciones séricas de todas las proteínas de fase aguda investigadas se mantuvieron estables durante el periodo de estudio y no difirieron significativamente de las concentraciones observadas a los 7 días de pre-inoculación (Figura 6).
PROTEÍNA C REACTIVA
La concentración sérica de CRP aumentó significativamente en los lechones inoculados solamente con el PRRSV y en los inoculados con ambos virus (Figura 6A) en comparación con los del grupo control (P < 0,05).
En los lechones del grupo PRRSV + swIAV, la concentración sérica media de CRP aumentó significativamente desde el día 2 post-inoculación hasta el final del estudio (en comparación con la concentración del día 0 y con los animales de control).
En el grupo de PRRSV, se observó una cinética diferente de la concentración sérica de CRP, ya que las concentraciones séricas medias de esta proteína fueron significativamente mayores a los 4, 10 y 14 DPI en comparación con el grupo control (P < 0,05).
LA CONCENTRACIÓN DE CRP AUMENTÓ SIGNIFICATIVAMENTE EN LOS GRUPOS PRRSV Y PRRSV + swIAV
HAPTOGLOBINA
La concentración sérica de Hp aumentó significativamente en los grupos PRRSV y PRRSV + swIAV en comparación con el grupo control (Figura 6B), siendo la dinámica de la concentración sérica de esta proteína de fase aguda similar en ambos grupos inoculados con PRRSV.
En los lechones de los grupos PRRSV + swIAV y PRRSV, las concentraciones séricas medias de Hp aumentaron significativamente desde el día 4 al día 14 post-inoculación (en comparación con las concentraciones séricas del día 0 y con el grupo control).
AMILOIDE SÉRICO A
Las concentraciones séricas de SAA aumentaron significativamente entre los días 2 y 4 post-inoculación en los grupos swIAV y PRRSV + swIAV en comparación con el grupo control (P < 0,05), pero a partir de los 5 dpi no hubo diferencias significativas en las concentraciones de esta proteína de fase aguda entre los grupos inoculados y el grupo control (P ≥ 0,05; Figura 6C).
NO HUBO DIFERENCIAS SIGNIFICATIVAS ENTRE LAS CONCENTRACIONES DE SAA ENTRE LOS GRUPOS INOCULADOS Y CONTROL A PARTIR DEL DÍA 5 POST-INOCULACIÓN
PIG-MAP
La concentración sérica de Pig-MAP se mantuvo sin cambios en comparación con su concentración previa a la inoculación (P ≥ 0,05) en los cerdos inoculados con swIAV y en los animales de control (Figura 6D).
En los lechones de los grupos de PRRSV y swIAV + PRRSV, se observaron aumentos significativos a partir del día 3 post-inoculación. La concentración sérica de Pig-MAP permaneció aumentada en ambos grupos hasta los 7 dpi en comparación con la concentración del día 0 y con las concentraciones en los cerdos de control (P < 0,05), no observándose diferencias en ambos grupos inoculados con PRRVS.
LESIONES PULMONARES
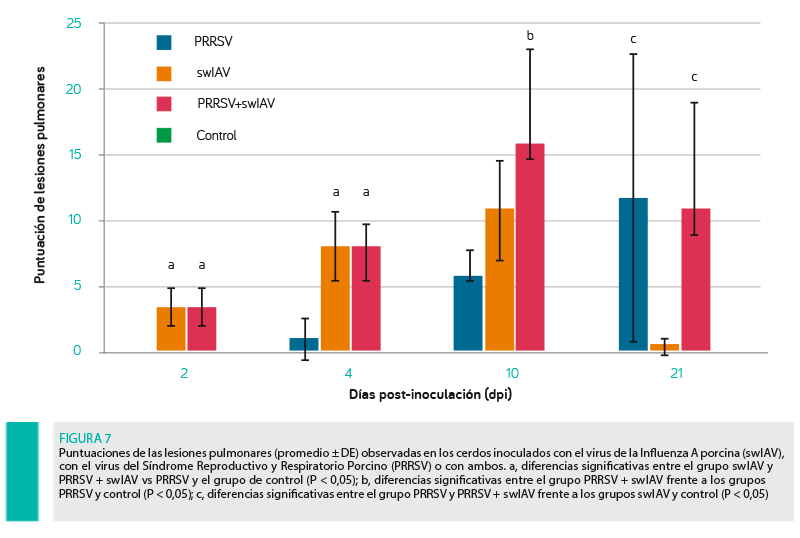
Se observaron lesiones pulmonares características de una infección viral de gravedad variable (puntuaciones de 1 a 15 en el grupo swIAV, 0 a 28 en el grupo PRRSV y 1,25 a 23,5 en el grupo swIAV + PRRSV) en todos los lechones inoculados con virus entre los días 2 y 10 post-inoculación. Los promedios de lesiones observadas en todos los grupos experimentales se presentan en la Figura 7.
A los 21 dpi, no se encontraron lesiones patológicas en 3 de los 5 lechones del grupo swIAV.
En el grupo PRRSV no se observaron lesiones a los 2 y 4 dpi mientras que a los 10 y 21 dpi se encontraron lesiones pulmonares típicas de la infección por PRRSV en todos los animales.
Los lechones del grupo control no mostraron lesiones patológicas.
A los 2 y 4 dpi, las puntuaciones de lesiones pulmonares observadas en los grupos swIAV y PRRSV + swIAV fueron significativamente mayores que las de los grupos PRRSV y control (P < 0,05).
A los 10 dpi, el promedio de puntuación de lesiones en el grupo PRRSV + swIAV fue mayor que en los grupos PRRSV y control (P < .05).
El promedio de la puntuación de lesiones difirió significativamente a los 21 dpi entre los grupos PRRSV y control (P < 0,05), así como en comparación con el grupo swIAV (P < 0,05).
CARGA VIRAL
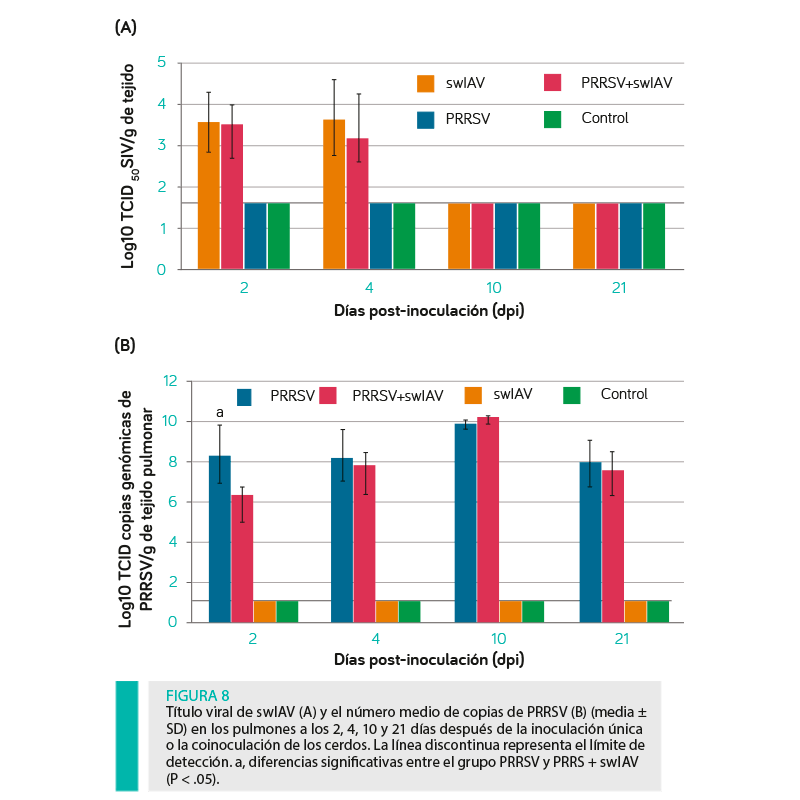
Las cargas virales en los pulmones a los 2, 4, 10 y 21 dpi se presentan en la Figura 8.
El ARN de swIAV se detectó en todas las muestras tomadas de los pulmones derechos de los cerdos inoculados con este virus a los 2 y 4 dpi. A los 10 y 21 dpi, no se detectó el swIAV en ninguno de los grupos. Los títulos medios de TCID50 del swIAV no difirieron significativamente entre los lechones inoculados solamente con este virus y los inoculados también con el PRRSV (P ≥ 0,05).
Se encontraron diferencias significativas entre el número medio de copias del PRRSV en los pulmones tomados de los cerdos de los grupos PRRSV y PRRSV + swIAV (P < 0,05) a los 2 dpi, siendo este valor significativamente mayor en los cerdos inoculados sólo con PRRSV. A los 4, 10 y 21 dpi, no se observaron diferencias significativas en la carga viral pulmonar de PRRSV (P ≥ .05).
El PRRSV se detectó en las muestras tomadas del pulmón derecho de todos los lechones inoculados solamente con PRRSV y aquellos inoculados también con swIAV desde el día 2 al 21 post-inoculación.
LA CARGA VIRAL DE PRRSV A LOS 2 DPI FUE SIGNIFICATIVAMENTE MAYOR EN EL GRUPO PRRVS EN COMPARACIÓN CON EL GRUPO PRRSV + swIAV
DISCUSIÓN
Los resultados de este estudio muestran que la infección simultánea con el PRRSV y el swIAV tiene un impacto limitado en el desarrollo clínico y la respuesta inmunitaria frente a estos virus. Para la mayoría de los parámetros estudiados, no se observaron diferencias significativas entre los lechones infectados de forma individual con PRRSV o con swIAV, o simultáneamente con ambos virus, incluyendo la puntuación de lesiones pulmonares.
No obstante, a pesar de que las puntuaciones clínicas no mostraron diferencias estadísticamente significativas, se observó un mayor número de lechones con fiebre persistente y de mayor duración en el grupo coinfectado.
Estudios previos sobre las coinfecciones de PRRSV y swIAV han arrojado resultados contradictorios en relación a su impacto en su evolución clínica7,6,13,17. En este caso, no se observó ningún retraso o descenso en la excreción de swIAV en el grupo coinfectado (PRRSV + swIAV) en comparación con el grupo swIAV.
En contraste, se evidenció un breve descenso de la replicación de PRRSV en los pulmones el día 2 post-inoculación en el grupo coinfectado (PRRSV + swIAV) en comparación con el grupo PRRSV (P < 0,05).
El impacto limitado de la coinfección en la evolución clínica, la carga viral y la excreción puede ser el resultado de las diferencias en la diana celular de los dos virus.
Los macrófagos diferenciados, concretamente, los macrófagos alveolares pulmonares (MAP) y los macrófagos intravasculares pulmonares, son la diana principal del PRRSV, aunque este virus también puede replicarse en menor medida, en células dendríticas y macrófagos derivados de monocitos presentes en la mayoría de los órganos22.
El swIAV infecta el epitelio del tracto respiratorio (bronquios, bronquiolos y alvéolos) y se replica ampliamente en los pulmones porcinos23.
El coronavirus respiratorio porcino (PRCV), que también se replica principalmente en el epitelio del tracto respiratorio inferior, interfiere fuertemente con la infección ocasionada por el swIAV, disminuyendo la replicación del swIAV en un 99%32. Por otro lado, cerdos coinfectados con PRRSV y PRCV muestran lesiones clínicas y pulmonares más severas, pero no se observó ningún impacto en el nivel de replicación ni en la eliminación del virus17.
Aunque un estudio anterior en el que se utilizó una línea celular recombinante susceptible de ser infectada por ambos virus, confirmó que los virus interferían entre sí24, en un hospedador natural sólo una pequeña población de células dendríticas y, en algunos casos, los neumocitos de tipo 1 pueden ser el objetivo común del PRRSV y del swIAV.
La interferencia del swIAV con la cantidad de PRRSV detectada a los 2 dpi podría ser el resultado de un aumento de la producción de interferón alfa (IFN-α) por parte de las células dendríticas plasmocitoides (pDC) y otros tipos de células en respuesta a la infección por el swIAV.
A los 2 dpi, cuando se detectó una disminución significativa de la replicación del PRRSV en los pulmones de los cerdos coinfectados (grupo PRRSV + swIAV), no se detectaron lesiones pulmonares visibles en el grupo PRRSV. Simultáneamente, la puntuación de lesiones en los grupos swIAV y PRRSV + swIAV eran comparables, lo que indica que durante este periodo de tiempo el swIAV fue la única causa de las lesiones patológicas en los pulmones.
Las diferencias en la puntuación de lesiones entre los grupos experimentales (infección individual o coinfección) observadas en este estudio probablemente están relacionadas con la dinámica de la infección por los virus (swIAV o PRRSV) y no con las interacciones que se producen entre ellos.
En el caso del swIAV, las lesiones pulmonares aparecieron antes y su disminución coincidió con el desarrollo más intenso de las lesiones pulmonares asociadas al PRRSV. El análisis del patrón de lesiones a lo largo del tiempo en el grupo coinfectado indicó que la puntuación reflejaba la dinámica aditiva de los grupos infectados únicamente con el swIAV o únicamente con el PRRSV. La puntuación de lesiones más alta se registró en el grupo coinfectado a los 10 dpi y superó significativamente la del grupo PRRSV. El promedio de la puntuación de lesiones en este momento también fue más alto, aunque no significativamente, en comparación con el grupo inoculado con swIAV.
Lo más probable es que el leve aumento de la gravedad de las lesiones clínicas y microscópicas, también señalado en estudios anteriores, sea un efecto del aumento del daño de la estructura pulmonar y de la inflamación en el caso de la coinfección13,17.
La infección por el swIAV provoca la necrosis de las células epiteliales, el aumento de la producción de mediadores proinflamatorios y la infiltración con células fagocíticas susceptibles a la infección por el PRRSV25,26.
Los signos respiratorios causados por el PRRSV también son el resultado de su acción sobre el pulmón y la interacción con el sistema inmunitario del hospedador. El virus provoca una neumonía intersticial e induce apoptosis mediada por TNF-α, también en los macrófagos transeúntes no infectados27.
Dado que este estudio se realizó en condiciones estrictamente controladas en una instalación BSL3, no refleja las condiciones de campo en las que pueden intervenir muchos factores. Concretamente, las diferentes cargas de infección bacteriana en el tracto respiratorio pueden interferir en el curso clínico y patológico de las coinfecciones víricas y contribuir al resultado.
Nuestros resultados también muestran que la coinfección con cepas locales de PRRSV y swIAV no afectó a la intensidad y cinética de las respuestas de fase aguda e inmunitaria. Los resultados de un estudio anterior determinaron que la respuesta de fase aguda era notablemente diferente entre cepas en términos de intensidad y duración28, pero la Hp fue el biomarcador más sensible para la infección por PRRSV. Además, la Hp y la CRP discriminaron entre los cerdos infectados y los de control.
Estos hallazgos coinciden con nuestros resultados, ya que el aumento significativo de Hp y CRP se observó sólo en los grupos PRRSV y PRRSV + swIAV. Además, se observaron diferencias significativas con respecto a la Hp entre los grupos de PRRSV y PRRSV + swIAV y el grupo de swIAV entre los días 4 y 14 post-infección.
No se observaron diferencias significativas en el nivel de proliferación de las PBMC (células mononucleares de sangre periférica) ni en la respuesta humoral frente a ambos patógenos entre los grupos infectados o coinfectados (P ≥ 0,05).
Aunque el PRRSV puede ser inmunosupresor y actuar de forma sinérgica con algunos patógenos, la coinfección simultánea de cerdos con el swIAV y el PRRSV no potenció la gravedad de los signos clínicos, las lesiones pulmonares, la respuesta inmunitaria y la replicación de ambos virus en el tracto respiratorio. La ausencia de sinergia entre ambos virus tras su replicación es beneficiosa para el hospedador, ya que no debería conducir a un empeoramiento de las alteraciones pulmonares y los signos clínicos, a pesar de la coinfección común en condiciones de campo.
1. Goka EA, Vallely PJ, Mutton KJ, et al. Single, dual and multiple respiratory virus infections and risk of hospitalization and mortality. Epidemiol Infect. 2015;143:37-47.
2. Martin ET, Kuypers J, Wald A, et al. Multiple versus single virus respiratory infections: viral load and clinical disease severity in hospitalized children. Influenza Other Respir Viruses. 2011;6:71-77.
3. Zhang G, Hu Y, Wang H, Zhang L, Bao Y, Zhou X. High incidence of multiple viral infections identified in upper respiratory tract infected children under three years of age in Shanghai, China. PLoS One. 2012;7(9):e44568.
4. Dorr APM, Gebreyes WA, Almond GW. Porcine reproductive and respiratory syndrome virus: age and management system disease modeling for pathogenic co-infection. J Swine Health Prod. 2007;15:258-264.
5. Opriessnig T, Gimenez-Lirola LG, Halbur PG. Polymicrobial respiratory disease in pigs. Animal Health Res Rev. 2011;12:133-148.
6. Van Reeth K, Nauwynck H, Pensaert M. Clinical effects of experimental dual infections with porcine reproductive and respiratory syndrome virus followed by swine influenza virus in conventional and colostrum-deprived pigs. J Vet Med B Infect Dis Vet Public Health. 2001;48:283-292.
7. Pol JM, van Leengoed LA, Stockhofe N, et al. Dual infections of PRRSV/influenza or PRRSV/Actinobacillus pleuropneumoniae in the respiratory tract. Vet Microbiol. 1997;55:259-264.
8. Jung K, Renukaradhya G, Alekseev KP, et al. Porcine reproductive and respiratory syndrome virus modifies innate immunity and alters disease outcome in pigs subsequently infected with porcine respiratory coronavirus: implications for respiratory viral co-infections. J Gen Virol. 2009;90:2713-2723.
9. Choi YK, Goyal SM, Joo HS. Retrospective analysis of etiologic agents associated with respiratory diseases in pigs. Can Vet J. 2003;44: 735-737.
10. Fablet C, Marois-Crehan C, Simon G, et al. Infectious agents associated with respiratory diseases in 125 farrow-to-finish pig herds: a cross-sectional study. Vet Microbiol. 2012;157:152-163.
11. Pinky L, Dobrovolny HM. Coinfections of the respiratory tract: viral competition for resources. PLoS One. 2016;11:e0155589.
12. Brand HK, de Groot R, Galama JMD, et al. Infection with multiple viruses is not associated with increased disease severity in children with bronchiolitis. Pediatr Pulmonol. 2012;47:393-400.
13. Kitikoon P, Vincent AL, Jones KR, et al. Vaccine efficacy and immune response to swine influenza virus challenge in pigs infected with porcine reproductive and respiratory syndrome virus at the time of SIV vaccination. Vet Microbiol. 2009;139:235-244.
14. Ouyang T, Zhang X, Liu X, et al. Co-infection of swine with porcine circovirus type 2 and other swine viruses. Viruses. 2019;11:185.
16. López-Soria S, Maldonado J, Riera P, et al. Selected swine viral pathogens in indoor pigs in Spain. Seroprevalence and farm-level characteristics. Transbound Emerg Dis. 2010;57:171-179.
17. Van Reeth K, Nauwynck H, Pensaert M. Dual infections of feeder pigs with porcine reproductive and respiratory syndrome virus followed by porcine respiratory coronavirus or swine influenza virus: a clinical and virological study. Vet Microbiol. 1996;48:325-335.
18. Mateu E, Diaz I. The challenge of PRRS immunology. Vet J. 2008;177:345-351.
19. Slomka MJ, Densham AL, Coward VJ, et al. Real time reverse transcription (RRT)-polymerase chain reaction (PCR) methods for detection of pandemic (H1N1) 2009 influenza virus and European swine influenza A virus infections in pigs. Influenza Other Respir Viruses. 2010;4:277-293.
20. Halbur PG, Paul PS, Frey ML, et al. Comparison of the pathogenicity of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet Pathol. 1995;32:648-660.
21. Pomorska-Mól M, Markowska-Daniel I. Interferon-γ secretion and proliferative responses of peripheral blood mononuclear cells after vaccination of pigs against Aujeszky’s disease in the presence of maternal immunity. FEMS Immunol Med Microbiol. 2010;58:405-411.
22. Singleton H, Graham SP, Bodman-Smith KB, et al. Establishing porcine monocyte-derived macrophage and dendritic cell systems for studying the interaction with PRRSV-1. Front Microbiol. 2016;7:832.
23. Van Reeth K, Nauwynck H. Proinflammatory cytokines and viral respiratory disease in pigs. Vet Res. 2010;31:187-213.
24. Provost C, Hamonic G, Gagnon CA, Meurens F. Dual infections of CD163 expressing NPTr epithelial cells with influenza A virus and PRRSV. Vet Microbiol. 2017;207:143-148.
25. Montoya M, Foni E, Solórzano A, et al. Expression dynamics of innate immunity in influenza virus-infected swine. Front Vet Sci. 2017;21:48.
26. Pomorska-Mol M, Markowska-Daniel I, Kwit K, et al. Immune and inflammatory response in pigs during acute influenza caused by H1N1 swine influenza virus. Arch Virol. 2014;159:2605-2614.
27. Choi C, Chae C. Expression of tumor necrosis factor-alpha is associated with apoptosis in lungs of pigs experimentally infected with porcine reproductive and respiratory syndrome virus. Res Vet Sci. 2002; 72:45-49.
28. Saco Y, Martínez-Lobo F, Cortey M, et al. C-reactive protein, haptoglobin and pig-major acute phase protein profiles of pigs infected experimentally by different isolates of porcine reproductive and respiratory syndrome virus. Vet Microbiol. 2016;183:9-15.





 El impacto de la coinfección en la gravedad y el resultado clínico no es del todo claro. Algunos estudios han demostrado que el resultado clínico de las coinfecciones víricas puede ser menos grave, o al menos no más grave, que la infección por un solo virus2,6,7,12,13. Por el contrario, otros estudios han constatado que la coinfección vírica agrava el curso clínico1,13.
El impacto de la coinfección en la gravedad y el resultado clínico no es del todo claro. Algunos estudios han demostrado que el resultado clínico de las coinfecciones víricas puede ser menos grave, o al menos no más grave, que la infección por un solo virus2,6,7,12,13. Por el contrario, otros estudios han constatado que la coinfección vírica agrava el curso clínico1,13.