Los rotavirus (RV) se clasifican en nueve especies o grupos (RVA-RVD y RVF-RVJ). Los RVA, RVB y RVC están bien reconocidos como agentes etiológicos de enfermedades entéricas en las explotaciones porcinas y han sido identificados en todos los países con una producción porcina relevante. Por su parte, los RVH sólo se han identificado en explotaciones porcinas de Japón y, más recientemente, de Brasil, EE.UU., Sudáfrica y Vietnam, pero todavía no en Europa. En nuestro estudio se investigó la presencia de RVH en 103 explotaciones porcinas españolas. Nueve explotaciones resultaron positivas y se obtuvo la secuencia nucleotídica completa en tres aislados y parcial en otro.
Las identidades de estas secuencias con respecto a las secuencias de RVH disponibles en la base de datos GenBank oscilaron entre el 69,4 % y el 93,7 %. Desde el punto de vista filogenético, todos los segmentos genómicos de los aislados españoles de RVH se agruparon estrechamente con otras cepas de RVH porcino, siendo mucho más lejana su relación con los RVH humanos y con la cepa de RVH de murciélago. Se trata de la primera descripción de RVH en granjas porcinas en Europa e incluye su caracterización mediante la secuenciación completa del genoma.
ROTAVIRUS
Los rotavirus (RV) son miembros de la familia Reoviridae y uno de los principales agentes causantes de gastroenteritis en humanos y animales en todo el mundo.
Su genoma consta de 11 segmentos de ARN de doble cadena que codifican seis proteínas estructurales (VP1-4, VP6 y VP7)y cinco proteínas no estructurales(NSP1-5)(Estes & Kapikian, 2007).
Según el Comité Internacional de Taxonomía de Virus, el género Rotavirus se divide en nueve grupos o especies antigénicamente distintas(RVA, RVB, RVC, RVD, RVF, RVG, RVH, RVI y RVJ) en función de la diversidad de la secuencia del gen que codifica la proteína de la cápside intermedia (VP6)(Matthijnssens et al., 2012; Mihalov-Kovács et al., 2015).
ROTAVIRUS PORCINOS
Las infecciones por RV son muy prevalentes en las explotaciones porcinas, frecuentemente ligadas a diarreas en lactación y post-destete, causando importantes pérdidas económicas para la industria porcina.
Los principales grupos de RV asociados a brotes de diarrea en el ganado porcino son[registrados] RVA, RVB y RVC.
Los RVA generalmente afectan a lechones de tres y cinco semanas de edad.
Los RVC son mucho más comunes en neonatos (<7 días de edad).
La detección de los RVB aumenta con la edad del animal, siendo más común identificarlos en diarreas de transición y cebo.
En cualquier caso, se ha comprobado que los tipos más prevalentes varían en función de la localización geográfica y también a lo largo del tiempo para una misma localización (Vlasova, Amimo, & Saif, 2017).
ROTAVIRUS H
En 1997, se describió en China un nuevo RV humano, denominado inicialmente como nuevo rotavirus de la diarrea del adulto, que no pertenecía a ningún grupo previamente establecido y que causó un brote de gastroenteritis entre adultos (Alam et al., 2007; Yang et al., 2004). Posteriormente, se identificó este nuevo rotavirus como RVH en base a la secuencia del gen que codifica para la proteína VP6(Matthijnssens et al., 2012).
En total, entre 1997 y 2002 se identificaron tres cepas humanas de RVH en Asia (ADRV-N y J19 en China y B219 en Bangladesh), así como una cepa porcina de RVH en Japón (SKA-1) (Jiang et al., 2008; Nagashima et al., 2008). Desde entonces, se han identificado RVH en cerdos de Japón (Suzuki & Inoue, 2018; Wakuda et al., 2011), EE.UU. (Marthaler et al., 2014), Brasil (Molinari, Lorenzetti, Otonel, Alfieri, & Alfieri, 2014), Sudáfrica (Nyaga et al., 2016) y Vietnam (Phan et al., 2016).
Aunque estos RVH se han detectado en las heces de cerdos con diarrea, las coinfecciones con otros patógenos son muy comunes y no se ha podido establecer de forma clara su papel en la etiología de la enfermedad entérica (Shepherd, Freeman, Culhane, & Marthaler, 2019). Más recientemente, se ha descrito la presencia de RVH en murciélagos de Camerún (Yinda et al., 2018).
Hasta donde sabemos, no hay informes previos de la presencia de RVH en cerdos en Europa. En este trabajo presentamos la primera detección y caracterización de RVH en cerdos con diarrea en granjas porcinas españolas, el principal productor de cerdos europeo.
MATERIALESY MÉTODOS
MUESTREO
El estudio se llevó a cabo entre 2017-2019 en 103 explotaciones comerciales porcinas españolas ubicadas en la mitad norte del país (22 provincias) con brotes de diarrea en los que se sospechaba una etiología viral.
Los brotes afectaron a lechones lactantes (<21 días) (26 granjas), etapa post-destete y transición (21-70 días) (11 granjas) o cerdos de engorde (>70 días) (33 granjas) y de cada uno de ellos, se enviaron entre 2 y 6 muestras de heces con fines diagnósticos al Departamento de Sanidad Animal de la Universidad de León (Tabla 1). La edad de los animales afectados no se conocía en 33 granjas.
EXTRACCIÓN Y DETECCIÓN DE ARN DE RVH
Para cada explotación, las muestras recibidas fueron mezcladas en una única muestra que se empleó para la extracción del ARN total utilizando un kit comercial (QIAamp Viral RNA Mini Kit, Qiagen) y siguiendo las instrucciones del fabricante.
A continuación, se realizó una PCR con transcripción inversa (RT-PCR) utilizando un par de cebadores de nuevo diseño basados en las secuencias del gen VP6 de las cepas de RVH porcinas disponibles en GenBank (Tabla 2), y que amplifican un fragmento de 1.240 nt.
Las reacciones de RT-PCR se llevaron a cabo con el kit Verso 1-Step RT-PCR ReddyMix (Thermo Scientific), siguiendo las recomendaciones del fabricante, con las siguientes condiciones:
1. Paso inicial: 50ºC durante 30 min y 95ºC durante 2 min.
2. Paso intermedio: 45 ciclos de 95ºC durante 20 s, 50ºC durante 30 s y 72ºC durante 1 min.
3. Paso final de extensión: 72ºC durante 10 min.
SECUENCIACIÓN DE MUESTRAS POSITIVAS A RVH
Para cada muestra identificada como positiva por RT-PCR a RVH, se llevó a cabo una nueva extracción del ARN total, utilizando en este caso TRIzol LS (Thermo Scientific). El ARN obtenido fue secuenciado directamente, sin necesidad de amplificación previa, en el Servicio de Bioinformática y Genómica (SGB) de la Universidad Autónoma de Barcelona (UAB).
La secuenciación de nueva generación (NGS) se llevó a cabo utilizando una plataforma Illumina Miseq y las secuencias de RVH se obtuvieron a partir de los resultados de la NGS aplicando el protocolo desarrollado por Cortey et al. (2019).
ANÁLISISFILOGENÉTICO
Cada segmento identificado fue confirmado mediante análisis BLAST de las secuencias ensambladas.
Las secuencias se alinearon utilizando CLUSTALW y las relaciones evolutivas entre las secuencias se examinaron mediante un análisis filogenético utilizando el método de máxima verosimilitud y el modelo de sustitución Tamura-Nei con el software MEGAX (Kumar, Stecher, Li, Knyaz, & Tamura, 2018).
Las secuencias obtenidas en este trabajo están depositadas en GenBank con los números de acceso MT644949-MT644992
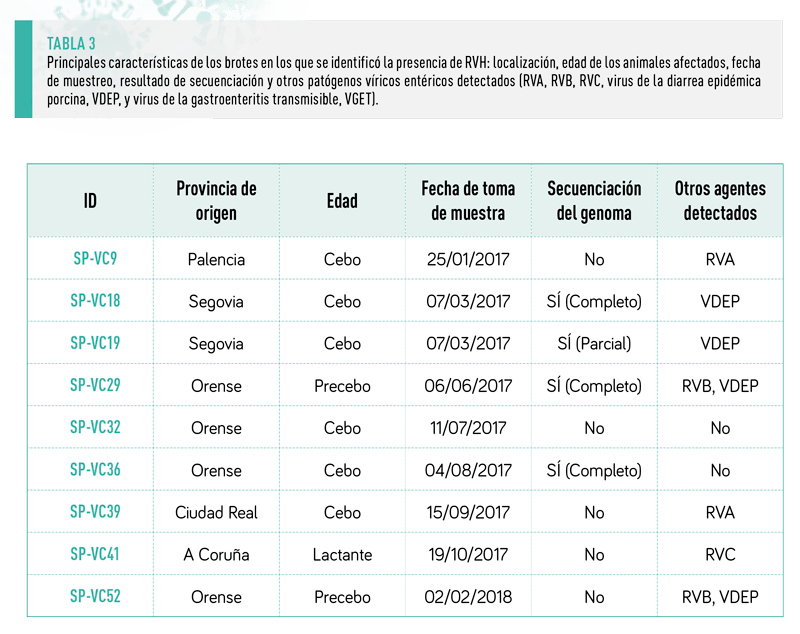
Se detectó RVH en nueve de las 103 explotaciones muestreadas (8,7 %), la mayoría de ellas con brote de diarrea en cerdos de engorde (6 muestras positivas de 33) o cerdos de transición (2 muestras positivas de 11). Sólo se detectó una muestra positiva en brotes de diarrea en lechones lactantes (1 de 26) (Tabla 3).
Este resultado concuerda con lo descrito en explotaciones porcinas de EE.UU. (15 % de muestras positivas), donde el riesgo de ser positivo a RVH, estimado por la Odds ratio, fue 5,9 mayor en los cerdos de más de 55 días en comparación con los lechones de entre 4 y 20 días (Marthaler et al., 2014).
En nuestra investigación, a pesar de las diferencias observadas, la proporción de brotes con detección positiva de RVH no difirió significativamente entre los grupos de edad cuando se comparó utilizando la prueba exacta de Fisher (p=0,139).
La eliminación de los anticuerpos maternos junto con la mezcla de lechones tras el destete puede explicar un mayor porcentaje de brotes positivos (18 %, 8 de 44) en cerdos postdestete (>21 días de edad) en comparación con los lechones lactantes (4%, 1 de 26).
EL RIESGO DE POSITIVIDAD A RVH ES MAYOR EN CERDOS TRANSICIÓN Y CEBO EN COMPARACIÓN CON LECHONES LACTANTES, LO QUE PODRÍA ATRIBUIRSE A LA DISMINUCIÓN DE LA INMUNIDAD MATERNAL Y LA MEZCLA DE ANIMALES TRAS EL DESTETE
COINFECCIÓN DE RVH CON OTROS VIRUS
RVH se detectó como coinfección con otros RV (A, B o C) en cinco brotes (55,5 %) y con el virus de la diarrea epidémica porcina (PEDV) en cuatro explotaciones (44,4 %)(Tabla 3).
Tan solo en dos de los brotes estudiados, ambos en cerdos de engorde, RVH fue el único agente etiológico viral identificado. No obstante, dado que no se investigó la presencia de agentes bacterianos causantes de diarrea como Brachyspira spp, Lawsonia intracellularis o Salmonella spp., no fue posible establecer la etiología definitiva en estas dos explotaciones.
ANÁLISIS FILOGENÉTICODE RVH
Se intentó llevar a cabo la secuenciación del genoma completo en todas las muestras positivas, obteniéndose el genoma completo (11 segmentos) en tres de ellas (SP-VC18, SP-VC29 y SP-VC36). En otra cepa (SP-VC19) se obtuvo la secuencia completa de los segmentos VP4, VP6, VP1, NSP1, NSP2, NSP4 y NSP5, mientras que fue parcial para VP7, VP2, VP3 y NSP3.
IDENTIDAD NUCLEOTÍDICA ENTRE LAS MUESTRAS POSITIVAS
La identidad nucleotídica entre las secuencias fue del:
82 %-100 % para VP7 83,2 %-100 % para VP4 86,1 %-100 % para VP6 87 %-100 % para VP1
84,4 %-100 % para VP2 81,6 %-100 % para VP3 84 %-100 % para NSP1 91 %-100 % para NSP2
83,5 %-100 % para NSP3 81,5 %-100 % para NSP4 92,3 %-100 % para NSP5
En concordancia con nuestros resultados, NSP5, NSP2 y VP1, que codifican proteínas directamente relacionadas con la replicación y el ensamblaje viral (fosfoproteína, NTPasa y ARN polimerasa dependiente de ARN, respectivamente), han sido propuestos como segmentos altamente conservados entre los RVH, mientras que VP3 y NSP4, que codifican la guaniltransferasa y la enterotoxina, respectivamente, presentan una mayor diversidad(Estes & Kapikian, 2007).
IDENTIDAD NUCLEOTÍDICA CEPAS DE RVH PORCINO DE ESPAÑA VS OTRAS CEPAS
Las cuatro secuencias de cepas de RVH porcino recuperadas en granjas españolas se compararon con las disponibles en GenBank, incluyendo secuencias parciales y completas del genoma de cepas de RVH porcino procedentes de Japón (n=11), EE.UU. (n=2), Brasil (n=3), Sudáfrica (n=1) y Vietnam (n=5), así como cepas de RVH humanas (n=3) y de murciélagos (n=1).
Al comparar todos los segmentos genómicos de las cepas españolas de RVH porcino con los genotipos propuestos de RVH porcino se observaron identidades de entre el 69,4 y el 93,7 %.
Las cepas españolas de RVH porcino estaban escasamente relacionadas con las cepas humanas de RVH (31,3-71,3 %), así como con la cepa de RVH de murciélago (15,9-68 %).
En concordancia, los árboles filogenéticos mostraron que las cepas españolas de RVH porcino estaban estrechamente relacionadas con las cepas de RVH de origen porcino de Japón, EE.UU., Brasil, Sudáfrica y Vietnam, y escasamente relacionadas con las cepas de RVH de origen humano de Bangladesh y China y que también eran distintas de una cepa de RVH identificada en murciélago en Camerún (Figuras 1 y 2).
Así pues, los aislados de RVH recuperados en diferentes hospedadores se agruparon en subgrupos claramente diferenciados (porcino, humano y murciélago), lo que sugiere la ausencia de eventos recientes de transmisión entre especies.
Esta observación se basa en un número muy limitado de cepas de RVH en las que se ha obtenido la secuencia completa del genoma, particularmente en el caso de aislados de origen humano y de murciélagos, siendo necesarios más estudios que aporten secuencias del genoma completo para determinar de forma concluyente las vías evolutivas de los RVH y su potencial zoonótico.
A PESAR DE QUE LOS ESTUDIOS NO MUESTRAN INDICIOS DE QUE EXISTA TRANSMISIÓN DE ROTAVIRUS ENTRE ESPECIES, SERÍA NECESARIO REALIZAR MÁS ESTUDIOS PARA EVALUAR SU POTENCIAL ZOONÓTICO
GENOTIPADO DE RVH BASADO EN SECUENCIAS GENÓMICAS COMPLETAS
Los rotavirus presentan una alta variabilidad, fruto de su genoma segmentado que favorece fenómenos de reasociación y reordenamiento. Esta variabilidad tiene repercusiones clínicas dado que la inmunidad frente a estos virus es dependiente del genotipo.
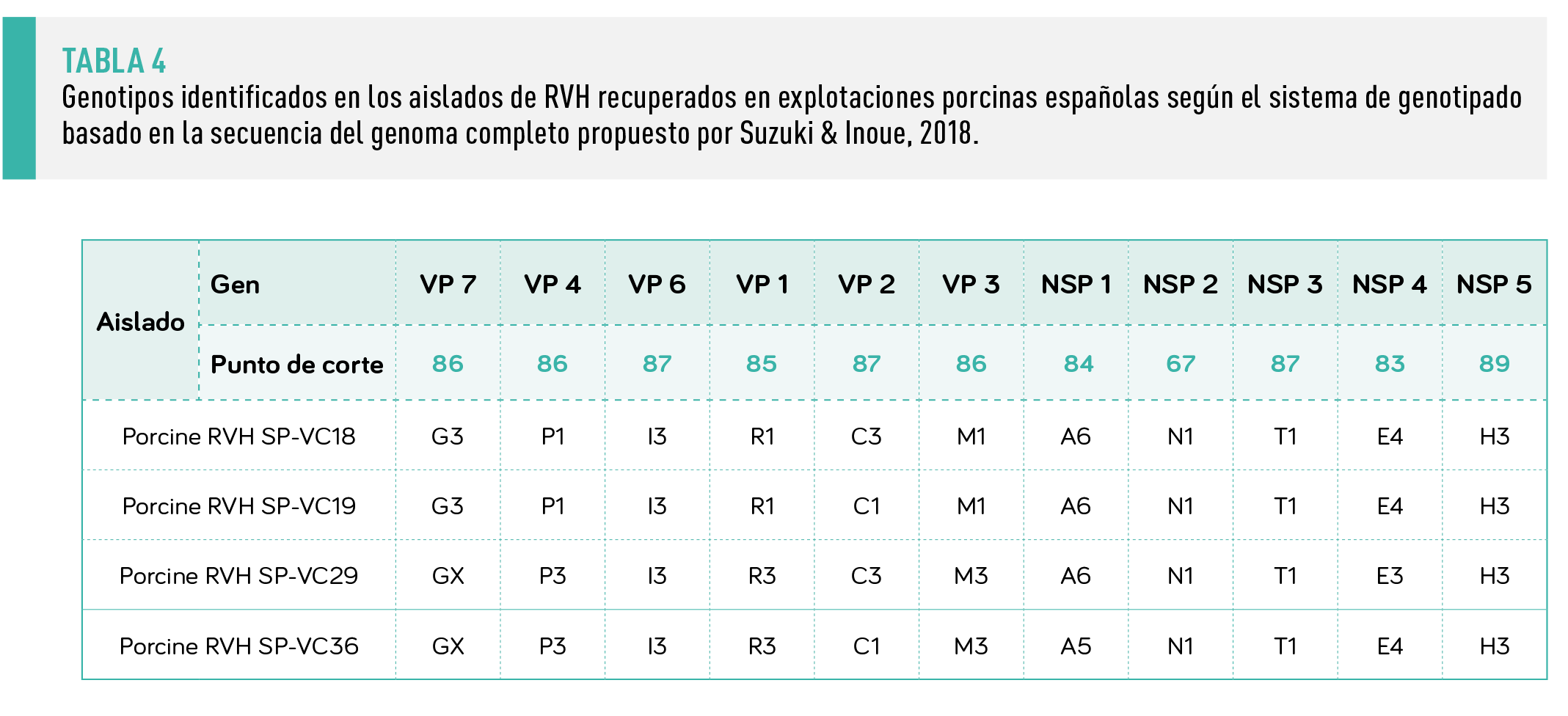
Se dispone de sistemas bien establecidos para el genotipado de RVA, pero no para otros grupos. No obstante, recientemente se ha propuesto un sistema de genotipado para los RVH basado en las secuencias genómicas completas(Suzuki & Inoue, 2018).
Teniendo en cuenta los valores de punto de corte recomendados, las cepas de RVH porcinas españolas mostraron cierta variabilidad, clasificándose en uno o dos genotipos diferentes para cada segmento genómico(Tabla 4 y Figura 1).
Se superaron valores de corte propuestos en el caso del segmento VP7 y las cepas SP-VC29 y SP-VC36 (genotipo X en la Figura 1 y Tabla 4). Sin embargo, este resultado debe tomarse con precaución debido al limitado número de secuencias disponibles.
Este hecho fue particularmente evidente al observar las secuencias correspondientes al segmento VP6(Figura 1), donde la agrupación del árbol parece apuntar a un único grupo muy diverso, mientras que, según los umbrales propuestos, el aislado SP-VC36 podría considerarse un nuevo genotipo.
SON NECESARIAS MÁS SECUENCIAS DE RVH PARA EL DESARROLLO ADECUADO Y LA REVISIÓN DE LOS UMBRALES EN EL SISTEMA DE GENOTIPADO PROPUESTO
CONCLUSIONES
En este trabajo se presentan las primeras secuencias genómicas de cuatro cepas de RVH porcinas (tres secuencias completas y una parcial) procedentes de España, siendo las primeras cepas de RVH identificadas en Europa.
Nuestros datos indican que los RVH están relativamente extendidos en la población porcina española, siendo identificados en casi el 9 % de los brotes de diarrea investigados. Además, la secuenciación completa del genoma demostró su utilidad en la caracterización de estos aislados, siendo de utilidad para el desarrollo de nuevas técnicas de diagnóstico que permitan la detección de RVH en las explotaciones porcinas y la vigilancia de esta infección en cerdos a nivel mundial.
Alam, M. M., Kobayashi, N., Ishino, M., Ahmed, M. S., Ahmed, M. U., Paul, S. K., … Naik, T. N. (2007). Genetic analysis of an ADRV-N-like novel rotavirus strain B219 detected in a sporadic case of adult diarrhea in Bangladesh. Archives of Virology, 152, 199-208. https://doi.org/10.1007/s00705-006-0831-y
Cortey, M., Díaz, I., Vidal, A., Martín-Valls, G., Franzo, G., Gómez De Nova, P. J., … Mateu, E. (2019). High levels of unreported intraspecific diversity among RNA viruses in faeces of neonatal piglets with diarrhoea. BMC Veterinary Research, 15, 1-13. https://doi.org/10.1186/s12917-019–2204-2
Estes, M., & Kapikian, A. (2007). Rotaviruses. En M. M. et al Knipe DM, Howley PM, Griffin DE, Lamb RA (Ed.), Fields Virology (5.a ed., pp. 1917-1974). Philadelphia: Lippincott Williams & Wilkins.
Jiang, S., Ji, S., Tang, Q., Cui, X., Yang, H., Kan, B., & Gao, S. (2008). Molecular characterization of a novel adult diarrhoea rotavirus strain J19 isolated in China and its significance for the evolution and origin of group B rotaviruses. Journal of General Virology, 89, 2622-2629. https://doi.org/10.1099/vir.0.2008/001933-0
Kumar, S., Stecher, G., Li, M., Knyaz, C., & Tamura, K. (2018). MEGA X: Molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution, 35, 1547-1549. https://doi.org/10.1093/molbev/msy096
Marthaler, D., Rossow, K., Culhane, M., Goyal, S., Collins, J., Matthijnssens, J., … Ciarlet, M. (2014). Widespread rotavirus H in domesticated pigs, United States. Emerging Infectious Diseases, 20, 1195-1198. https://doi.org/10.3201/eid2007.140034
Matthijnssens, J., Otto, P. H., Ciarlet, M., Desselberger, U., van Ranst, M., & Johne, R. (2012). VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Archives of Virology, 157,1177-1182. https://doi.org/10.1007/s00705-012-1273-3
Mihalov-Kovács, E., Gellért, Á., Marton, S., Farkas, S. L., Fehér, E., Oldal, M., … Bányai, K. (2015). Candidate new Rotavirus species in sheltered Dogs, Hungary. Emerging Infectious Diseases, 21, 660-663. https://doi.org/10.3201/eid2104.141370
Molinari, B. L. D.T, Lorenzetti, E., Otonel, R. A. A., Alfieri, A. F., & Alfieri, A. A. (2014). Rotavirus Detected in Piglets with. Emerging Infectious Diseases, 20, 1019-1022. https://doi.org/http://dx.doi.org/10.3201/eid2006.130776
Nagashima, S., Kobayashi, N., Ishino, M., Alam, M. M., Ahmed, M. U., Shyamal Kumar Paul, S. K., … Wang, Y. H. (2008). Whole Genomic Characterization of a Human Rotavirus Strain B219 Belonging to a Novel Group of the Genus Rotavirus Shigeo. Journal of Medical Virology, 80, 2023-2033. https://doi.org/10.1002/jmv
Nyaga, M. M., Peenze, I., Potgieter, C. A., Seheri, L. M., Page, N. A., Yinda, C. K., … Mphahlele, M. J. (2016). Complete genome analyses of the first porcine rotavirus group H identified from a South African pig does not provide evidence for recent interspecies transmission events. Infection, Genetics and Evolution, 38, 1-7. https://doi.org/10.1016/j.meegid.2015.11.032
Phan, M. V. T., Anh, P. H., Cuong, N. Van, Munnink, B. B. O., Hoek, L. Vander, My, P. T., … Cotten, M. (2016). Unbiased whole-genome deep sequencing of human and porcine stool samples reveals circulation ofmultiple groups of rotaviruses and a putative zoonotic infection. Virus Evolution, 2, 1-15. https://doi.org/10.1093/ve/vew027
Shepherd, F. K., Freeman, M. J., Culhane, M. R., & Marthaler, D. G. (2019). Diseases of swine. En J. J.
Zimmerman, L. A. Karriker, A. Ramirez, K. J. Schwartz, G. W. Stevenson, & J. Zhang (Eds.), Diseases of swine (11th ed., pp. 715-727). Hoboken: Wiley-Blackwell. https://doi.org/10.1017/CBO9781107415324.004
Suzuki, T., & Inoue, D. (2018). Full genome-based genotyping system for rotavirus H and detection of potential gene recombination in nonstructural protein 3 between porcine rotavirus H and rotavirus C. Journal of General Virology, 99, 1582-1589. https://doi.org/10.1099/jgv.0.001162
Vlasova, A. N., Amimo, J. O., & Saif, L. J. (2017). Porcine rotaviruses: Epidemiology, immune responses and control strategies. Viruses, 9, 1-27. https://doi.org/10.3390/v9030048
Wakuda, M., Ide, T., Sasaki, J., Komoto, S., Ishii, J., Sanekata, T., & Taniguchi, K. (2011). Porcine rotavirus closely related to novel group of human rotaviruses. Emerging Infectious Diseases, 17,1491-1493. https://doi.org/10.3201/eid1708.101466
Yang, H., Makeyev, E. V., Kang, Z., Ji, S., Bamford, D. H., & Van Dijk, A. A. (2004). Cloning and sequence analysis of dsRNA segments 5, 6 and 7 of a novel non-group A, B, C adult rotavirus that caused an outbreak of gastroenteritis in China. Virus Research, 106, 15-26. https://doi.org/10.1016/j.virusres.2004.05.011
Yinda, C. K., Ghogomu, S. M., Conceição-Neto, N., Beller, L., Deboutte, W., Vanhulle, E., … Matthijnssens, J. (2018). Cameroonian fruit bats harbor divergent viruses, including rotavirus H, bastroviruses, and picobirnaviruses using an alternative genetic code. Virus Evolution, 4, 1-15. https://doi.org/10.1093/ve/vey008


 Las infecciones por RV son muy prevalentes en las explotaciones porcinas, frecuentemente ligadas a diarreas en lactación y post-destete, causando importantes pérdidas económicas para la industria porcina.
Las infecciones por RV son muy prevalentes en las explotaciones porcinas, frecuentemente ligadas a diarreas en lactación y post-destete, causando importantes pérdidas económicas para la industria porcina.